
Effects of a N-Maleimide-derivatized Phosphatidylethanolamine on the Architecture and Properties of Lipid Bilayers †
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
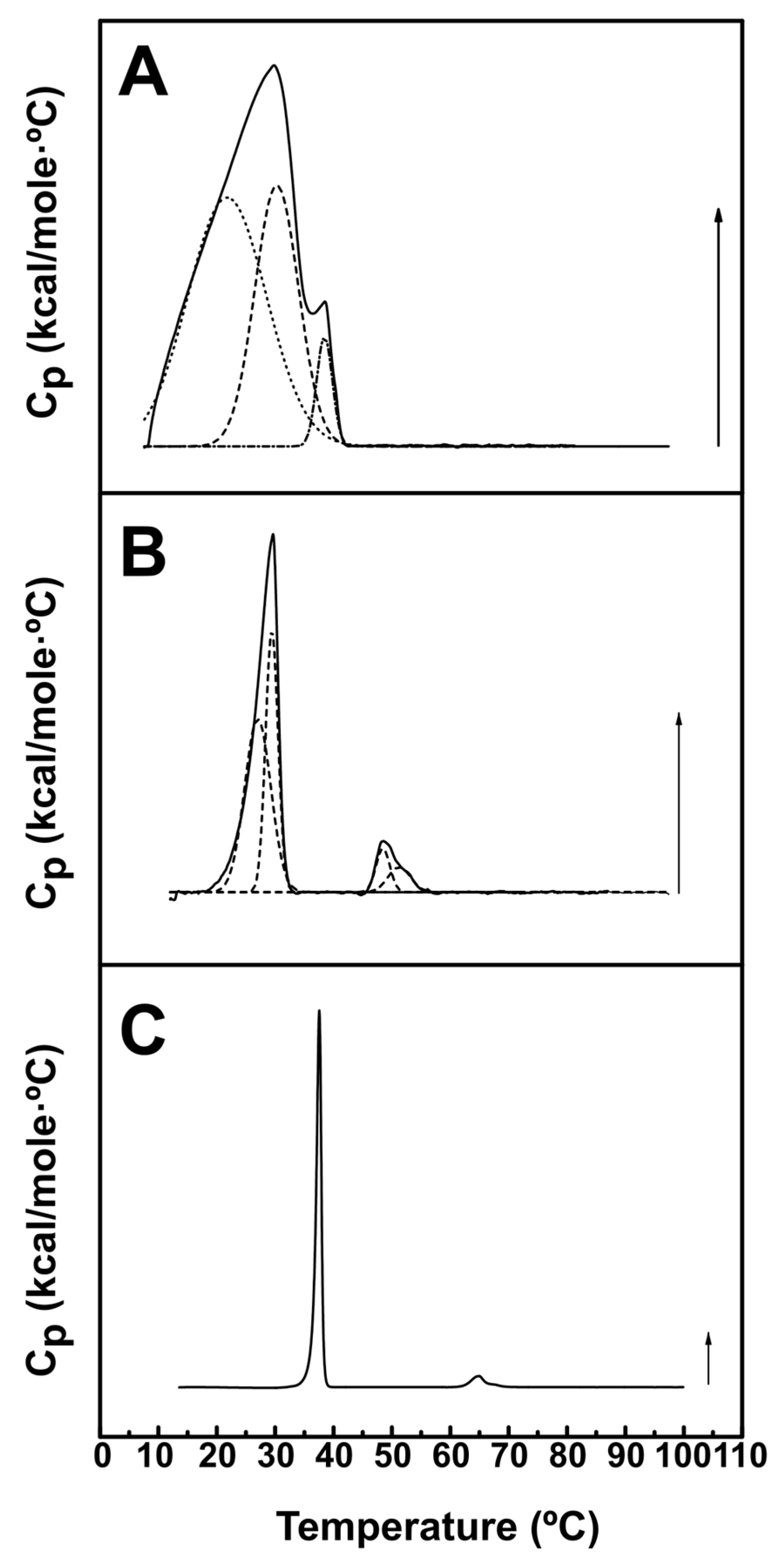
2.1. Calorimetric Studies
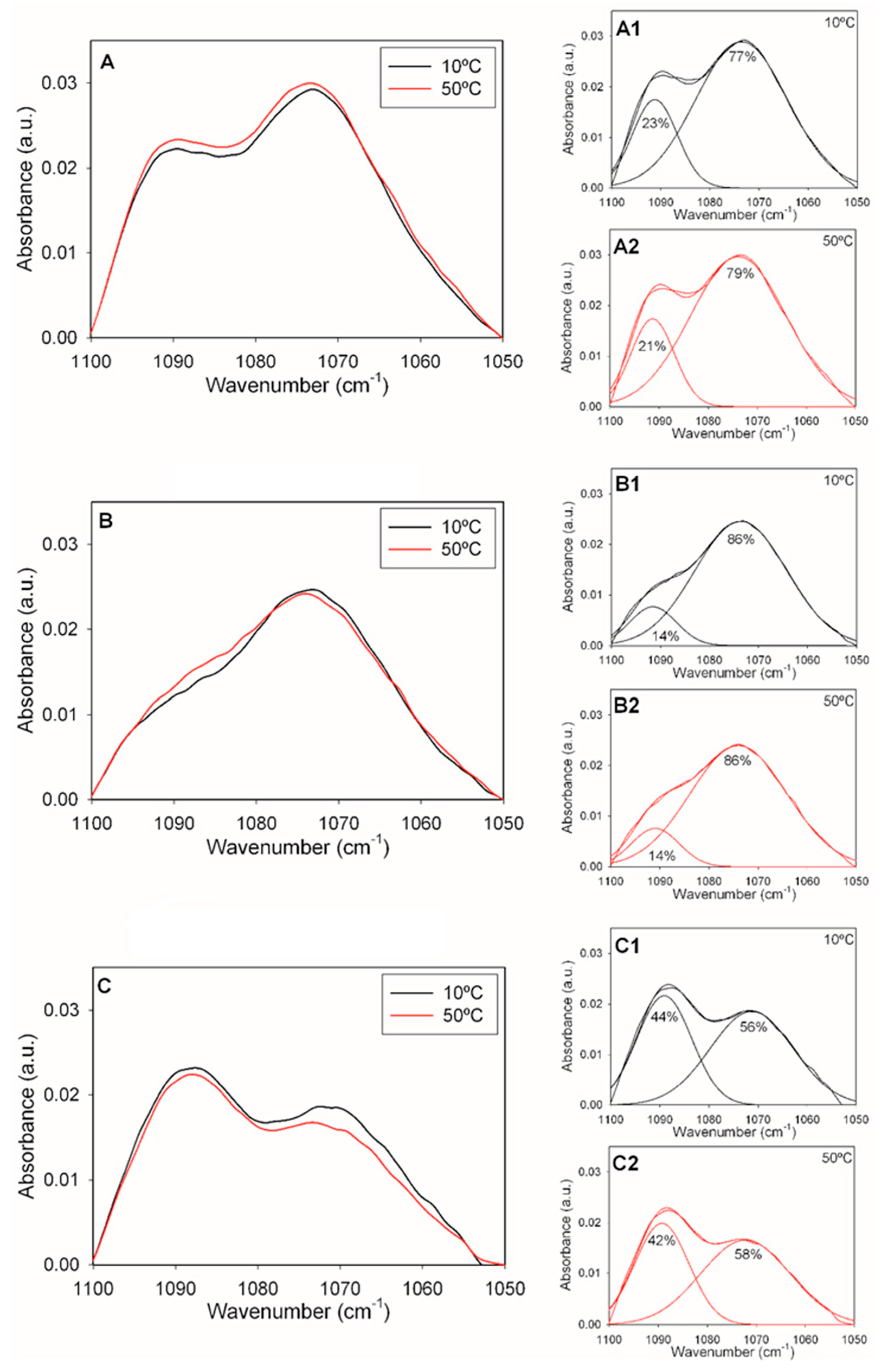
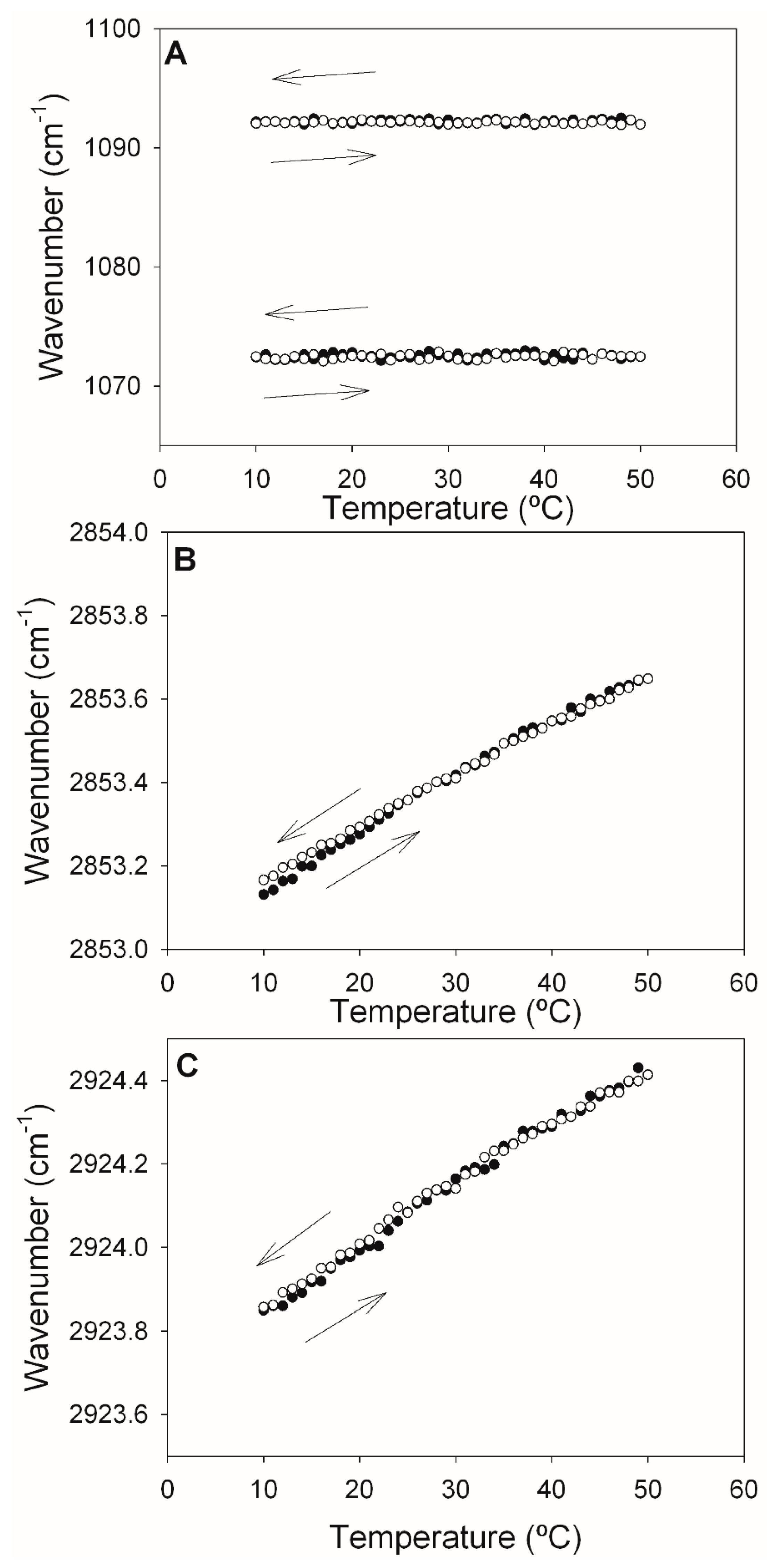
2.2. Infrared Spectroscopy
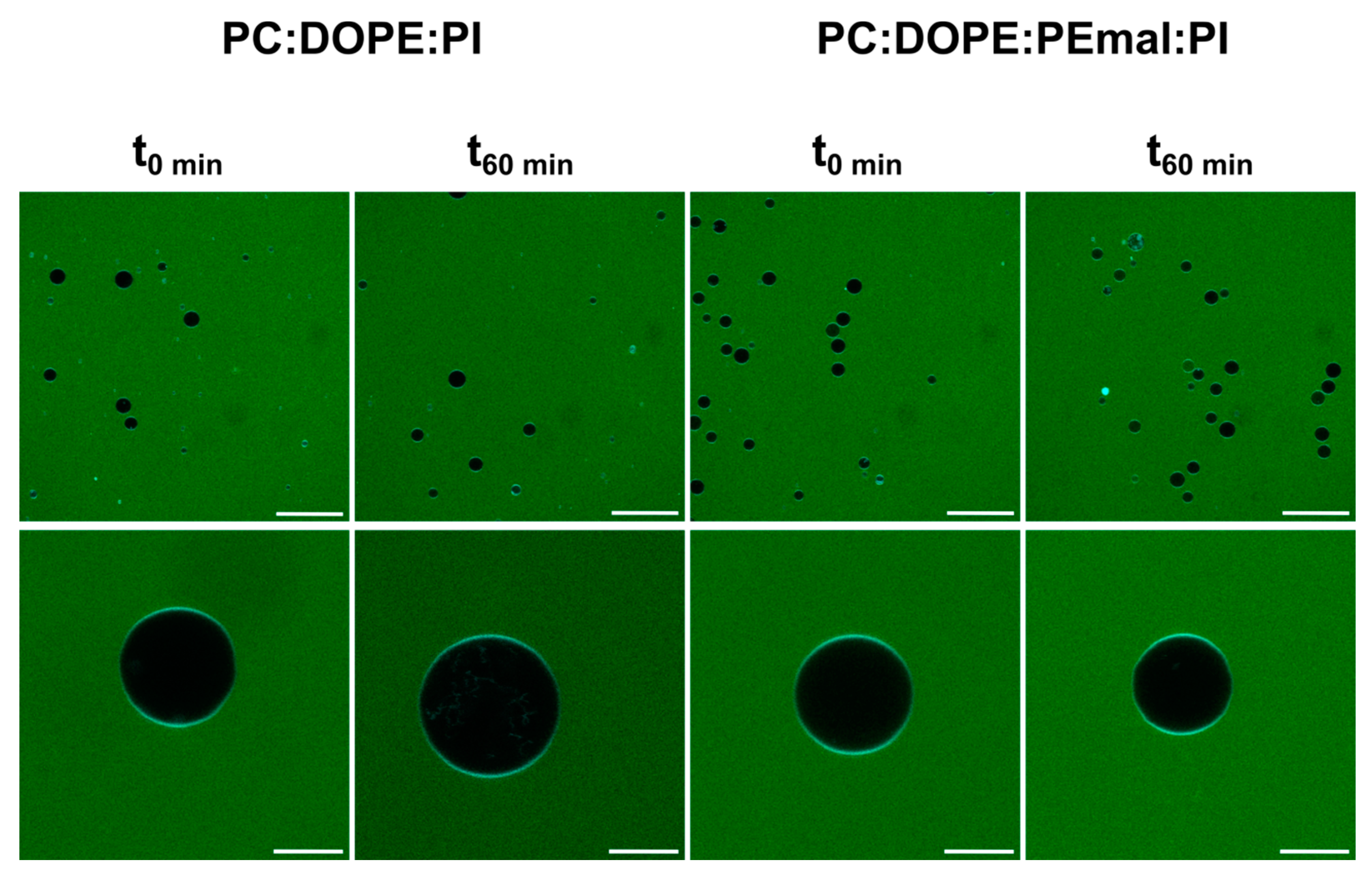
2.3. Fluorescence Studies
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Vesicle Preparation
4.3. Differential Scanning Calorimetry (DSC)
4.4. IR Spectroscopy
4.5. Vesicle Leakage Assessment (Fluorescence Spectroscopy)
4.6. Vesicle Permeability Assessment (Fluorescence Microscopy)
4.7. Membrane Fluidity Measurements (Laurdan and DPH)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Elliott, J.T.; Prestwich, G.D. Maleimide-Functionalized Lipids That Anchor Polypeptides to Lipid Bilayers and Membranes. Bioconjug. Chem. 2000, 11, 832–841. [Google Scholar] [CrossRef]
- Schuy, S.; Treutlein, B.; Pietuch, A.; Janshoff, A. In Situ Synthesis of Lipopeptides as Versatile Receptors for the Specific Binding of Nanoparticles and Liposomes to Solid-Supported Membranes. Small 2008, 4, 970–981. [Google Scholar] [CrossRef]
- Takeshige, K.; Baba, M.; Tsuboi, S.; Noda, T.; Ohsumi, Y. Autophagy in Yeast Demonstrated with Proteinase-Deficient Mutants and Conditions for Its Induction. J. Cell Biol. 1992, 119, 301–312. [Google Scholar] [CrossRef]
- Nishimura, T.; Tooze, S.A. Emerging Roles of ATG Proteins and Membrane Lipids in Autophagosome Formation. Cell Discov. 2020, 6, 32. [Google Scholar] [CrossRef]
- Melia, T.J.; Lystad, A.H.; Simonsen, A. Autophagosome Biogenesis: From Membrane Growth to Closure. J. Cell Biol. 2020, 219, e202002085. [Google Scholar] [CrossRef]
- Nakatogawa, H.; Ichimura, Y.; Ohsumi, Y. Atg8, a Ubiquitin-like Protein Required for Autophagosome Formation, Mediates Membrane Tethering and Hemifusion. Cell 2007, 130, 165–178. [Google Scholar] [CrossRef]
- Weidberg, H.; Shpilka, T.; Shvets, E.; Abada, A.; Shimron, F.; Elazar, Z. LC3 and GATE-16 N Termini Mediate Membrane Fusion Processes Required for Autophagosome Biogenesis. Dev. Cell 2011, 20, 444–454. [Google Scholar] [CrossRef]
- Landajuela, A.; Hervás, J.H.; Antón, Z.; Montes, L.R.; Gil, D.; Valle, M.; Rodriguez, J.F.; Goñi, F.M.; Alonso, A. Lipid Geometry and Bilayer Curvature Modulate LC3/GABARAP-Mediated Model Autophagosomal Elongation. Biophys. J. 2016, 110, 411–422. [Google Scholar] [CrossRef]
- Iriondo, M.N.; Etxaniz, A.; Varela, Y.R.; Ballesteros, U.; Hervás, J.H.; Montes, L.R.; Goñi, F.M.; Alonso, A. LC3 Subfamily in Cardiolipin-Mediated Mitophagy: A Comparison of the LC3A, LC3B and LC3C Homologs. Autophagy 2022, 18, 2985–3003. [Google Scholar] [CrossRef]
- Ballesteros, U.; Etxaniz, A.; Iriondo, M.N.; Varela, Y.R.; Lázaro, M.; Viguera, A.R.; Montes, L.R.; Valle, M.; Goñi, F.M.; Alonso, A. Autophagy Protein LC3C Binding to Phospholipid and Interaction with Lipid Membranes. Int. J. Biol. Macromol. 2022, 212, 432–441. [Google Scholar] [CrossRef]
- Wang, M.T.; Ma, R.; Wu, C.X. Membrane buckling and the determination of Gaussian curvature modulus. Phys. Rev. E 2023, 108, 034802. [Google Scholar] [CrossRef]
- Marsh, D. Intrinsic curvature in normal and inverted lipid structures and in membranes. Biophys. J. 1996, 70, 2248–2255. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A Ubiquitin-like System Mediates Protein Lipidation. Nature 2000, 408, 488–492. [Google Scholar] [CrossRef]
- Xie, Z.; Nair, U.; Klionsky, D.J. Atg8 Controls Phagophore Expansion during Autophagosome Formation. Mol. Biol. Cell 2008, 19, 3290–3298. [Google Scholar] [CrossRef]
- Maruyama, T.; Alam, J.M.; Fukuda, T.; Kageyama, S.; Kirisako, H.; Ishii, Y.; Shimada, I.; Ohsumi, Y.; Komatsu, M.; Kanki, T.; et al. Membrane Perturbation by Lipidated Atg8 Underlies Autophagosome Biogenesis. Nat. Struct. Mol. Biol. 2021, 28, 583–593. [Google Scholar] [CrossRef]
- Ladbrooke, B.D.; Williams, R.M.; Chapman, D. Studies on Lecithin-Cholesterol-Water Interactions by Differential Scanning Calorimetry and X-Ray Diffraction. Biochim. Biophys. Acta 1968, 150, 333–340. [Google Scholar] [CrossRef]
- Sot, J.; Aranda, F.J.; Collado, M.I.; Goñi, F.M.; Alonso, A. Different Effects of Long- and Short-Chain Ceramides on the Gel-Fluid and Lamellar-Hexagonal Transitions of Phospholipids: A Calorimetric, NMR, and X-Ray Diffraction Study. Biophys. J. 2005, 88, 3368–3380. [Google Scholar] [CrossRef]
- Nele, V.; D’Aria, F.; Campani, V.; Silvestri, T.; Biondi, M.; Giancola, C.; De Rosa, G. Unravelling the role of lipid composition on liposome-protein interactions. J Liposome Res. 2023, 1–9. [Google Scholar] [CrossRef]
- Arrondo, J.L.R.; Goñi, F.M.; Macarulla, J.M. Infrared Spectroscopy of Phosphatidylcholines in Aqueous Suspension. A Study of the Phosphate Group Vibrations. Biochim. Biophys. Acta 1984, 794, 165–168. [Google Scholar] [CrossRef]
- Ellens, H.; Bentz, J.; Szoka, F.C. H+- and Ca2+-Induced Fusion and Destabilization of Liposomes. Biochemistry 1985, 24, 3099–3106. [Google Scholar] [CrossRef]
- Sot, J.; García-Arribas, A.B.; Abad, B.; Arranz, S.; Portune, K.; Andrade, F.; Martín-Nieto, A.; Velasco, O.; Arana, E.; Tueros, I.; et al. Erythrocyte Membrane Nanomechanical Rigidity Is Decreased in Obese Patients. Int. J. Mol. Sci. 2022, 23, 1920. [Google Scholar] [CrossRef]
- Płaczkiewicz, J.; Gieczewska, K.; Musiałowski, M.; Adamczyk-Popławska, M.; Bącal, P.; Kwiatek, A. Availability of iron ions impacts physicochemical properties and proteome of outer membrane vesicles released by Neisseria gonorrhoeae. Sci. Rep. 2023, 13, 18733. [Google Scholar] [CrossRef]
- Gómez-Murcia, V.; Torrecillas, A.; de Godos, A.M.; Corbalán-García, S.; Gómez-Fernández, J.C. Both idebenone and idebenol are localized near the lipid-water interface of the membrane and increase its fluidity. Biochim. Biophys. Acta. 2016, 1858, 1071–1081. [Google Scholar] [CrossRef]
- Mayer, L.D.D.; Hope, M.J.J.; Cullis, P.R.R. Vesicles of Variable Sizes Produced by a Rapid Extrusion Procedure. Biochim. Biophys. Acta-Biomembr. 1986, 858, 161–168. [Google Scholar] [CrossRef]
- Fiske, C.H.; Subbarow, Y. The Colorimetric Determination of Phosphorous. J. Biol. Chem. 1925, 66, 375–400. [Google Scholar] [CrossRef]
- Dimitrov, D.S.; Angelova, M.I. Lipid Swelling and Liposome Formation Mediated by Electric Fields. Bioelectrochem. Bioenerg. 1988, 19, 323–336. [Google Scholar] [CrossRef]
- Varela, Y.R.; González-Ramírez, E.J.; Iriondo, M.N.; Ballesteros, U.; Etxaniz, A.; Montes, L.R.; Goñi, F.M.; Alonso, A. Lipids in Mitochondrial Macroautophagy: Phase Behavior of Bilayers Containing Cardiolipin and Ceramide. Int. J. Mol. Sci. 2023, 24, 5080. [Google Scholar] [CrossRef]
- Oñate-Garzón, J.; Ausili, A.; Manrique-Moreno, M.; Torrecillas, A.; Ar-anda, F.J.; Patiño, E.; Gomez-Fernández, J.C. The increase in positively charged residues in cecropin D-like Galleria mellonella favors its interaction with mem-brane models that imitate bacterial membranes. Arch. Biochem. Biophys. 2017, 629, 54–62. [Google Scholar] [CrossRef]
- Arrondo, J.L.R.; Goñi, F.M. Structure and Dynamics of Membrane Proteins as Studied by Infrared Spectroscopy. Prog. Biophys. Mol. Biol. 1999, 72, 367–405. [Google Scholar] [CrossRef]
- de la Arada, I.; Gonzalez-Ramirez, E.J.; Alonso, A.; Goni, F.M.; Arrondo, J.-L.R. Exploring Polar Headgroup Interactions between Sphingomyelin and Ceramide with Infrared Spectroscopy. Sci. Rep. 2020, 10, 17606. [Google Scholar] [CrossRef]
- Cameron, D.G.; Casal, H.L.; Mantsch, H.H.; Boulanger, Y.; Smith, I.C. The Thermotropic Behavior of Dipalmitoyl Phosphatidylcholine Bilayers. A Fourier Transform Infrared Study of Specifically Labeled Lipids. Biophys. J. 1981, 35, 1–16. [Google Scholar] [CrossRef]
- Goñi, F.M.; Villar, A.V.; Nieva, J.L.; Alonso, A. Interaction of Phospholipases C and Sphingomyelinase with Liposomes. Methods Enzymol. 2003, 372, 3–19. [Google Scholar] [CrossRef]
- Parasassi, T.; Gratton, E.; Yu, W.M.; Wilson, P.; Levi, M. Two-Photon Fluorescence Microscopy of Laurdan Generalized Polarization Domains in Model and Natural Membranes. Biophys. J. 1997, 72, 2413–2429. [Google Scholar] [CrossRef]
- Sanchez, S.A.; Tricerri, M.A.; Gratton, E. Laurdan generalized polarization fluctuations measures membrane packing micro-heterogeneity in vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 7314–7319. [Google Scholar] [CrossRef]
- Lentz, B.R. Use of Fluorescent Probes to Monitor Molecular Order and Motions within Liposome Bilayers. Chem. Phys. Lipids 1993, 64, 99–116. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Tm (°C) | T1/2 (°C) | ΔH (kcal/mol) |
|---|---|---|---|
| DPPC:DOPEmal (70:30) | 31.9 ± 0.8 | 10.1 ± 0.2 | 6.8 ± 1.7 |
| DPPC:DOPE (70:30) | 34.5 ± 0.7 | 8.9 ± 0.4 | 7.2 ± 0.7 |
| DPPC | 41.9 | 0.7 | 9.0 |
| Sample | Transition | Tm (°C) | T1/2 (°C) | ΔH (kcal/mol) |
|---|---|---|---|---|
| DEPE:PE-mal (70:30) | 1 | |||
| a | 15.7 ± 3.6 | 14.6 ± 3.7 | 6.6 ± 4.2 | |
| b | 25.8 ± 2.8 | 10.5 ± 1.9 | 4.7 ± 2.9 | |
| c | 33.7 ± 2.9 | 4.7 ± 2.9 | 1.4 ± 1.6 | |
| DEPE:DOPE (70:30) | 1 | |||
| a | 26.1 ± 1.3 | 6.8 ± 2.1 | 1.0 ± 0.5 | |
| b | 29.2 ± 0.4 | 3.0 ± 0.7 | 0.7 ± 0.3 | |
| 2 | ||||
| a | 48.5 ± 0.1 | 3.2 ± 0.6 | 0.14 ± 0.06 | |
| b | 51.6 ± 0.5 | 4.5 ± 0.0 | 0.11 ± 0.03 | |
| DEPE | 1 | 37.6 ± 0.1 | 0.9 ± 0.0 | 10.3 ± 0.4 |
| 2 | 64.8 ± 0.1 | 3.1 ± 0.1 | 0.8 ± 0.0 |
| % Leakage (t180–t0) | 20 °C | 37 °C | 50 °C |
|---|---|---|---|
| ePC:DOPE:PI | 7.2 ± 3.0 | 7.8 ± 1.8 | 6.3 ± 5.9 |
| ePC:DOPE:DOPEmal:PI | 6.1 ± 4.3 | 8.1 ± 3.1 | 11.4 ± 2.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballesteros, U.; González-Ramirez, E.J.; de la Arada, I.; Sot, J.; Etxaniz, A.; Goñi, F.M.; Alonso, A.; Montes, L.R. Effects of a N-Maleimide-derivatized Phosphatidylethanolamine on the Architecture and Properties of Lipid Bilayers. Int. J. Mol. Sci. 2023, 24, 16570. https://doi.org/10.3390/ijms242316570
Ballesteros U, González-Ramirez EJ, de la Arada I, Sot J, Etxaniz A, Goñi FM, Alonso A, Montes LR. Effects of a N-Maleimide-derivatized Phosphatidylethanolamine on the Architecture and Properties of Lipid Bilayers. International Journal of Molecular Sciences. 2023; 24(23):16570. https://doi.org/10.3390/ijms242316570
Chicago/Turabian StyleBallesteros, Uxue, Emilio J. González-Ramirez, Igor de la Arada, Jesús Sot, Asier Etxaniz, Félix M. Goñi, Alicia Alonso, and Lidia Ruth Montes. 2023. "Effects of a N-Maleimide-derivatized Phosphatidylethanolamine on the Architecture and Properties of Lipid Bilayers" International Journal of Molecular Sciences 24, no. 23: 16570. https://doi.org/10.3390/ijms242316570