Response of the Bacterial Community and Antibiotic Resistance in Overnight Stagnant Water from a Municipal Pipeline

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Turbidity and Residual Chlorine
2.3. Heterotrophic Bacteria Plate Count (HPC)
2.4. DNA Extraction and High-Throughput Sequencing
2.5. ARB Calculation of the Antibiotic Resistance Rate
2.5.1. Antibiotic Exposure
2.5.2. Quantitative Assessment of Antibiotic Resistance
2.6. Statistical Analysis
3. Results and Discussion
3.1. Turbidity, HPC and Residual Chlorine
3.2. Response of the Bacterial Community
3.2.1. Bacterial Phyla
3.2.2. Bacterial Genera
3.3. Antibiotic Resistance
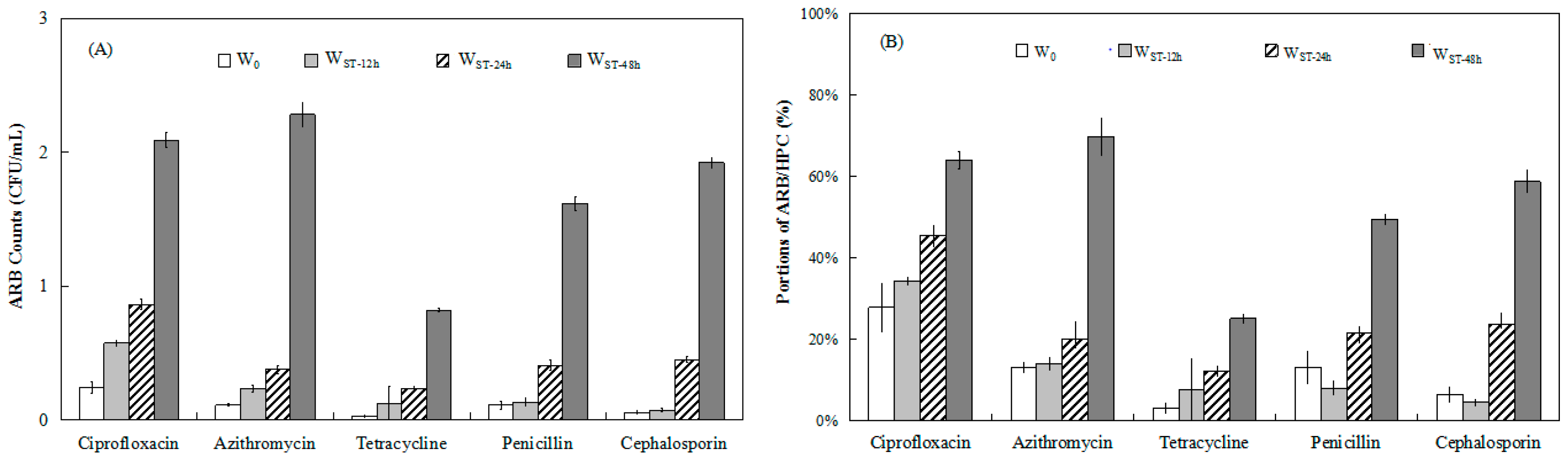
3.3.1. ARB Distribution
3.3.2. Antibiotic Resistance Level
3.4. Redundancy Analysis (RDA) of Bacterial Communities and Antibiotic Resistance
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zietz, B.P.; de Vergara, J.D.; Dunkelberg, H. Copper concentrations in tap water and possible effects on infant’s health-results of a study in Lower Saxony, Germany. Environ. Res. 2003, 92, 129–138. [Google Scholar] [CrossRef]
- Zietz, B.P.; Lass, J.; Suchenwirth, R. Assessment and management of tap water lead contamination in Lower Saxony, Germany. Int. J. Environ. Health Res. 2007, 17, 407–418. [Google Scholar] [CrossRef]
- Haider, T.; Haider, M.; Wruss, W.; Sommer, R.; Kundi, M. Lead in drinking water of Vienna in comparison to other European countries and accordance with recent guidelines. Int. J. Hyg. Environ. Health 2002, 205, 399–403. [Google Scholar] [CrossRef]
- Zlatanović, L.; van der Hoek, J.P.; Vreeburg, J.H.G. An experimental study on the influence of water stagnation and temperature change on water quality in a full-scale domestic drinking water system. Water Res. 2017, 123, 761–772. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Diversity and antibiotic resistance in Pseudomonas spp. from drinking water. Sci. Total Environ. 2012, 426, 366–374. [Google Scholar] [CrossRef]
- Xi, C.; Zhang, Y.; Marrs, C.F.; Ye, W.; Simon, C.; Foxman, B.; Nriagu, J. Prevalence of antibiotic resistance in drinking water treatment and distribution systems. Appl. Environ. Microb. 2009, 75, 5714–5718. [Google Scholar] [CrossRef] [Green Version]
- Narciso-da-Rocha, C.; Vaz-Moreira, I.; Svensson-Stadler, L.; Moore, E.R.; Manaia, C.M. Diversity and antibiotic resistance of Acinetobacter spp. in water from the source to the tap. Appl. Microbiol. Biot. 2013, 97, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Fang, W.; Hu, J.; Ong, S.L. Effects of phosphorus on biofilm disinfections in model drinking water distribution systems. J. Water Health 2010, 8, 446–454. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zeng, S.; He, M.; Gu, A.Z. Water disinfection byproducts induce antibiotic resistance-role of environmental pollutants in resistance phenomena. Environ. Sci. Technol. 2016, 50, 3193–3201. [Google Scholar] [CrossRef]
- Jiang, L.; Hu, X.; Xu, T.; Zhang, H.; Sheng, D.; Yin, D. Prevalence of antibiotic resistance genes and their relationship with antibiotics in the Huangpu River and the drinking water sources, Shanghai, China. Sci. Total Environ. 2013, 458–460, 267–272. [Google Scholar] [CrossRef]
- Talukdar, P.K.; Rahman, M.; Nabi, A.; Islam, Z.; Hoque, M.M.; Endtz, H.P.; Islam, M.A. Antimicrobial resistance, virulence factors and genetic diversity of Escherichia coli isolates from household water supply in Dhaka, Bangladesh. PLoS ONE 2013, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.L.; Wang, L.; Xu, M.Y.; Zhou, H.; Wang, S.L.; Wang, Y.J.; Bai, M. Selective antibiotic resistance genes in multiphase samples during biofilm growth in a simulated drinking water distribution system: Occurrence, correlation and low-pressure ultraviolet removal. Sci. Total Environ. 2019, 649, 146–155. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Health of the People’s Republic of China, Standardization Administration of China, GB/T5750-2006 Standard examination methods for drinking water. Available online: http://www.nhc.gov.cn/wjw/pgw/201212/33644.shtml (accessed on 14 March 2020).
- Huang, J.J.; Hu, H.Y.; Lu, S.Q.; Li, Y.; Tang, F.; Lu, Y.; Wei, B. Monitoring and evaluation of antibiotic-resistant bacteria at a municipal wastewater treatment plant in China. Environ. Int. 2012, 42, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Delean, A.; Munson, P.J.; Rodbar, D. Simultaneous analysis of families of sigmoidal curves: Application to bioassay, radioligand assay, and physiological dose-response curves. Am. J. Physiol. Endoc. M. 1978, 235, 97–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karin, L.; Nico, B.; Yingying, W.; Thomas, E.; Frederik, H. Overnight stagnation of drinking water in household taps induces microbial growth and changes in community composition. Water Res. 2010, 44, 4868–4877. [Google Scholar]
- Kahlisch, L.; Henne, K.; Groebe, L.; Draheim, J.; Höfle, M.G.; Brettar, I. Molecular analysis of the bacterial drinking water community with respect to live/dead status. Water Sci. Technol. 2010, 61, 9–14. [Google Scholar] [CrossRef]
- Jia, S.; Shi, P.; Hu, Q.; Li, B.; Zhang, T.; Zhang, X.X. Bacterial community shift drives antibiotic resistance promotion during drinking water chlorination. Environ. Sci. Technol. 2015, 49, 12271–12279. [Google Scholar] [CrossRef]
- Vaz-Moreira, I.; Nunes, O.C.; Manaia, C.M. Ubiquitous and persistent, Proteobacteria and other Gram-negative bacteria in drinking water. Sci. Total Environ. 2017, 586, 1141–1149. [Google Scholar] [CrossRef]
- Henne, K.; Kahlisch, L.; Brettar, I.; Höfle, M.G. Analysis of structure and composition of bacterial core communities in mature drinking water biofilms and bulk water of a citywide network in Germany. Appl. Environ. Microb. 2012, 78, 3530–3538. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.F.; Shi, B.Y.; Bai, Y.H.; Wang, D.S. Bacterial community of biofilms developed under different water supply conditions in a distribution system. Sci. Total Environ. 2014, 472, 99–107. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, J.; Mi, Z.; Xie, S.; Chen, C.; Zhang, X. Biofilm bacterial communities in urban drinking water distribution systems transporting waters with different purification strategies. Appl. Microbiol. Biot. 2015, 99, 1947–1955. [Google Scholar] [CrossRef] [PubMed]
- Martiny, A.C.; Albrechtsen, H.J.; Arvin, E.; Molin, S. Identification of bacteria in biofilm and bulk water samples from a nonchlorinated model drinking water distribution system. Appl. Environ. Microb. 2005, 71, 8611–8617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef] [PubMed]
- Prest, E.I.; Hammes, F.; Loosdrecht, M.; Vrouwenvelder, J.S. Biological stability of drinking water: Controlling factors, methods, and challenges. Front. Microbiol. 2016, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Hoefel, D.; Monis, P.T.; Grooby, W.L.; Andrews, S.; Saint, C.P. Profiling bacterial survival through a water treatment process and subsequent distribution system. J. Appl. Microbiol. 2005, 99, 175. [Google Scholar] [CrossRef]
- Zhang, M.L.; Liu, W.J.; Li, C.P.; Li, Y.X.; Gu, J.N. Effect of water switch on the bacterial community structure of finished water in a drinking water treatment plant. China. Environ. Sci. 2015, 35, 2517–2522. [Google Scholar]
- Navarro-Noya, Y.E.; Suárez-Arriaga, M.C.; Rojas-Valdes, A.; Montoya-Ciriaco, N.M.; Gómez-Acata, S.; Fernández-Luqueño, F.; Dendooven, L. Pyrosequencing analysis of the bacterial community in drinking water wells. Microb. Ecol. 2013, 66, 19–29. [Google Scholar] [CrossRef]
- Chao, Y.; Mao, Y.; Wang, Z.; Zhang, T. Diversity and functions of bacterial community in drinking water biofilms revealed by high-throughput sequencing. Sci. Rep. 2015, 5, 10044. [Google Scholar] [CrossRef]
- Douterelo, I.; Sharpe, R.L.; Boxall, J.B. Influence of hydraulic regimes on bacterial community structure and composition in an experimental drinking water distribution system. Water Res. 2013, 47, 503–516. [Google Scholar] [CrossRef]
- Kalyuzhnaya, M.G.; Bowerman, S.; Lara, J.C.; Lidstrom, M.E.; Chistoserdova, L. Methylotenera mobilis gen. nov., sp. nov., an obligately methylamine-utilizing bacterium within the family Methylophilaceae. Int. J. Syst. Evol. Microbiol. 2006, 56, 2819–2823. [Google Scholar] [CrossRef] [Green Version]
- Jiao, S.; Liu, Z.; Lin, Y.; Jun, Y.; Chen, W.; Wei, G. Bacterial communities in oil contaminated soils: Biogeography and co-occurrence patterns. Soil Biol. Biochem. 2016, 98, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, W.; Chen, J.; Qi, W.; Wang, F.; Zhou, Y.Y. Impact of biofilm formation and detachment on the transmission of bacterial antibiotic resistance in drinking water distribution systems. Chemosphere 2018, 203, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.C.; Kolvenbach, B.A.; Nunes, O.C.; Corvini, P.F.X. Biodegradation of antibiotics: The new resistance determinants–part II. New Biotechno. 2020, 54, 13–27. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, R.; Yunesian, M.; Nasseri, S.; Gholami, M.; Jalilzadeh, E.; Shoeibi, S.; Mesdaghinia, A. Occurrence and fate of most prescribed antibiotics in different water environments of Tehra, Iran. Sci. Total Environ. 2018, 619–620, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Catherine, L.T.; Philip, H.; Eilis, C.B.; Charlotte, K.C.; Viivi, H.A.; Yuiko, T.; James, S. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar]
- Reinthaler, F.F.; Posch, J.; Feierl, G.; Wüst, G.; Haas, D.; Ruckenbauer, G.; Mascher, F.; Marth, E. Antibiotic resistance of E. coli in sewage and sludge. Water Res. 2003, 37, 1690. [Google Scholar] [CrossRef]
- Lu, S.Q.; Li, Y.; Huang, J.J.; Wei, B.; Hu, H.Y. Antibiotic resistance of bacteria to 6 antibiotics in secondary effluents of municipal wastewater treatment plants, China. Environ. Sci. 2011, 32, 3419–3424. [Google Scholar]
- Su, H.C.; Liu, Y.S.; Bai, Y.H.; Pan, C.G.; Chen, J.; He, Y.L.; Ying, G.G. Persistence of antibiotic resistance genes and bacterial community changes in drinking water treatment system: From drinking water source to tap water. Sci. Total Environ. 2018, 616-617, 453–461. [Google Scholar] [CrossRef]
- Benotti, M.J.; Trenholm, R.A.; Vanderford, B.J.; Holady, J.C.; Stanford, B.D.; Snyder, S.A. Pharmaceuticals and Endocrine Disrupting Compounds in U.S. Drinking Water. Environ. Sci. Technol. 2009, 43, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.R.; Wang, Q.J.; Mo, C.H.; Li, Y.W.; Gao, P.; Tai, Y.P.; Zhang, Y.; Ruan, Z.L.; Xu, J.W. Determination of four fluoroquinolone antibiotics in tap water in Guangzhou and Macao. Environ. Pollut. 2010, 158, 2350–2358. [Google Scholar]
- Cheng, W.; Jiang, L.; Lu, N.; Ma, L.; Sun, X.; Luo, Y.; Lin, K.; Cui, C. Development of a method for trace level determination of antibiotics in drinking water sources by high performance liquid chromatography-tandem mass spectrometry. Anal. Methods. 2015, 7, 1777–1787. [Google Scholar] [CrossRef]
- Shen, Q.H.; Ji, X.L.; Fu, S.J.; Liu, Y.Y.; Li, L. Preliminary studies on the pollution levels of antibiotic and antibiotic resistance genes in Huangpu River, China. Ecol. Environ. Sci. 2012, 10, 1717–1723. [Google Scholar]
- Berendonk, T.U.; Manaia, C.M.; Merlin, C.; Fatta-Kassinos, D.; Cytryn, E.; Walsh, F.; Bürgmann, H.; Sørum, H.; Norström, M.; Pons, M.N. Tackling antibiotic resistance: The environmental framework. Nat. Rev. Microbiol. 2015, 13, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Cernohorska, L.; Votava, M. Determination of minimal regrowth concentration (MRC) in clinical isolates of various biofilm-forning bacteria. Folia microbial. 2004, 49, 75–78. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Xu, M.; Xu, S.; Zhang, L.; Lin, K.; Zhang, L.; Bai, M.; Zhang, C.; Zhou, H. Response of the Bacterial Community and Antibiotic Resistance in Overnight Stagnant Water from a Municipal Pipeline. Int. J. Environ. Res. Public Health 2020, 17, 1995. https://doi.org/10.3390/ijerph17061995
Zhang M, Xu M, Xu S, Zhang L, Lin K, Zhang L, Bai M, Zhang C, Zhou H. Response of the Bacterial Community and Antibiotic Resistance in Overnight Stagnant Water from a Municipal Pipeline. International Journal of Environmental Research and Public Health. 2020; 17(6):1995. https://doi.org/10.3390/ijerph17061995
Chicago/Turabian StyleZhang, Minglu, Mengyao Xu, Shaofeng Xu, Lingyue Zhang, Kaizong Lin, Lei Zhang, Miao Bai, Can Zhang, and He Zhou. 2020. "Response of the Bacterial Community and Antibiotic Resistance in Overnight Stagnant Water from a Municipal Pipeline" International Journal of Environmental Research and Public Health 17, no. 6: 1995. https://doi.org/10.3390/ijerph17061995