

Castor Plant Adaptation to Salinity Stress during Early Seedling Stage by Physiological and Transcriptomic Methods
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Stress Treatments
2.2. Determination of Plant Biomass
2.3. Determination of Malondialdehyde (MDA) Content
2.4. Determination of Soluble Sugar and Proline Content
2.5. Transcriptome Analysis
2.5.1. RNA Extraction and Sequencing and Analysis of Sequence Data
2.5.2. Quantitative Reverse Transcription–Polymerase Chain Reaction (qRT-PCR) Analysis
2.6. Statistical Analysis
3. Results
3.1. Growth and Content of Malondialdehyde, Soluble Sugars, and Proline under Salt Stress
3.2. Assembly and Transcriptome Quality Assessment
3.3. Gene Expression and Differentially Expressed Gene Analysis
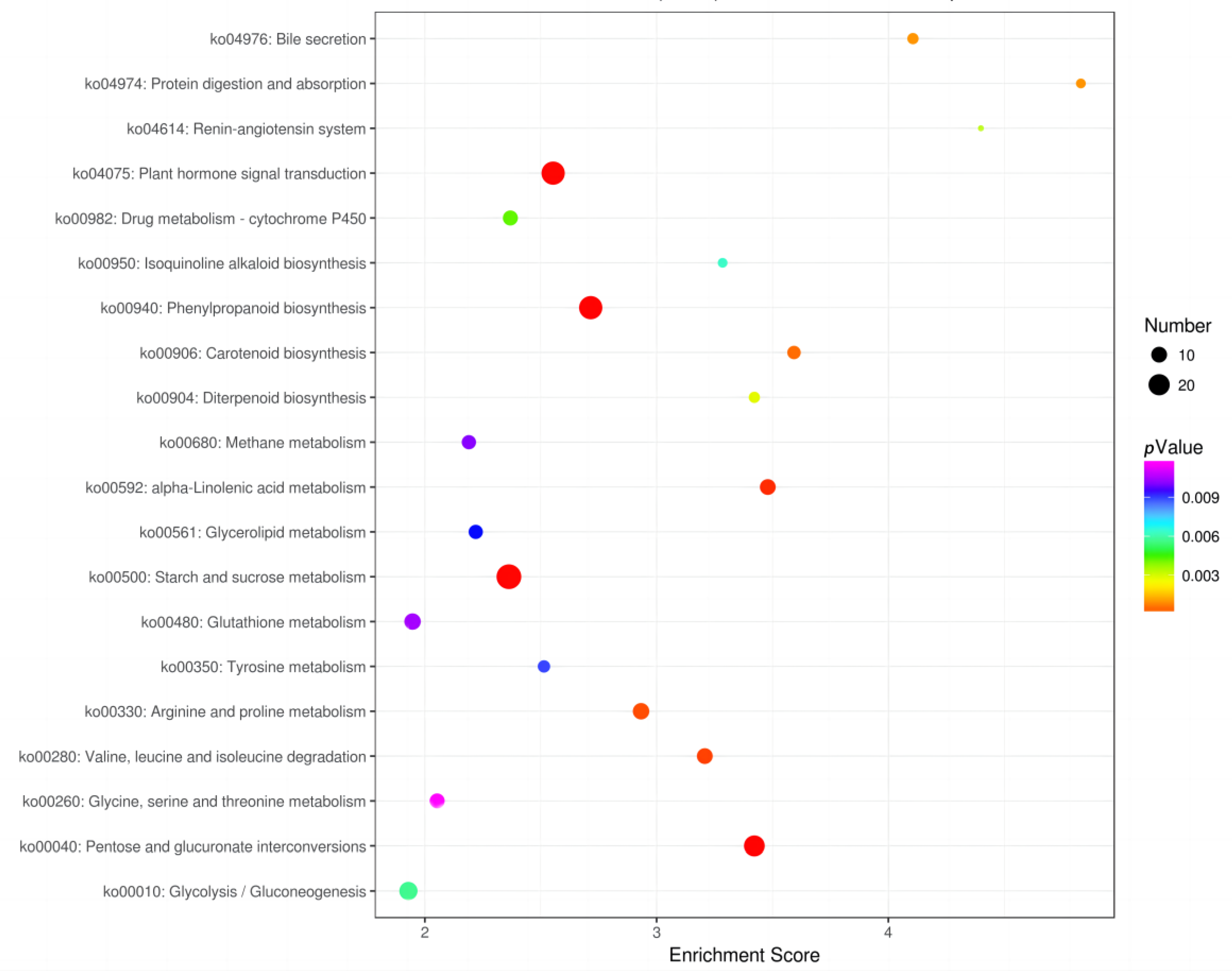
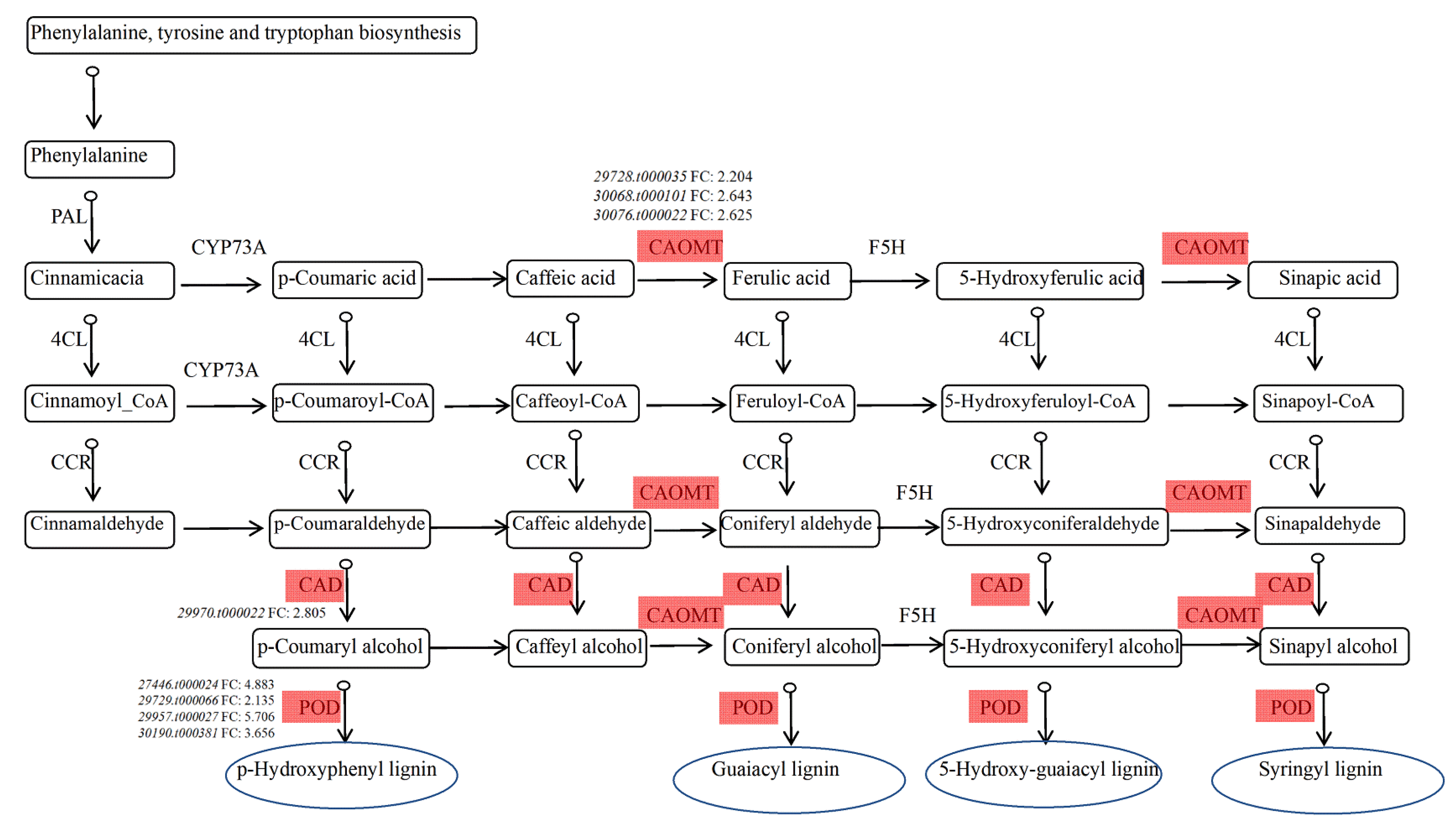
3.4. Functional Annotation and Enrichment Analysis
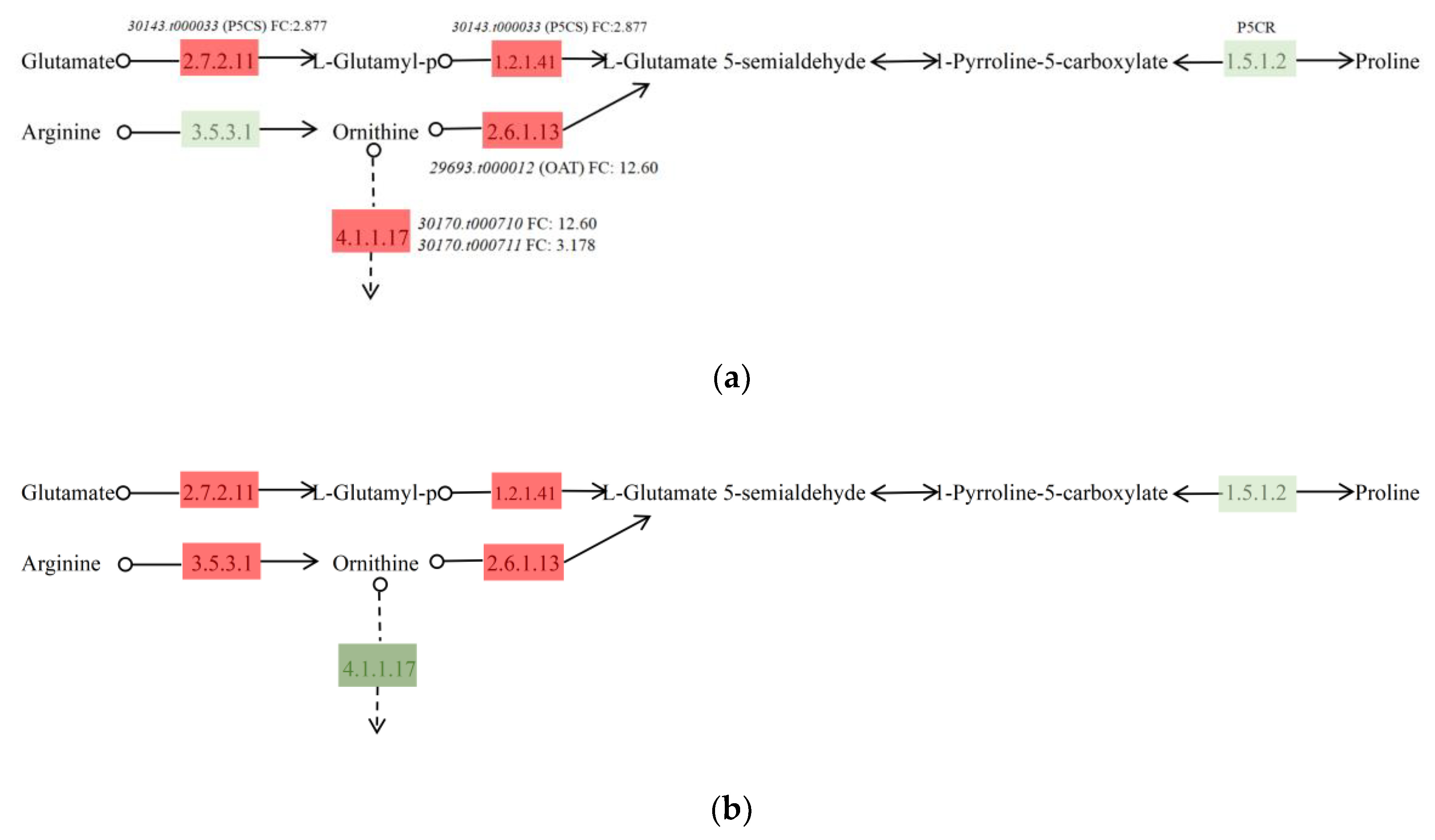
3.5. TF Responses to Salt Stress
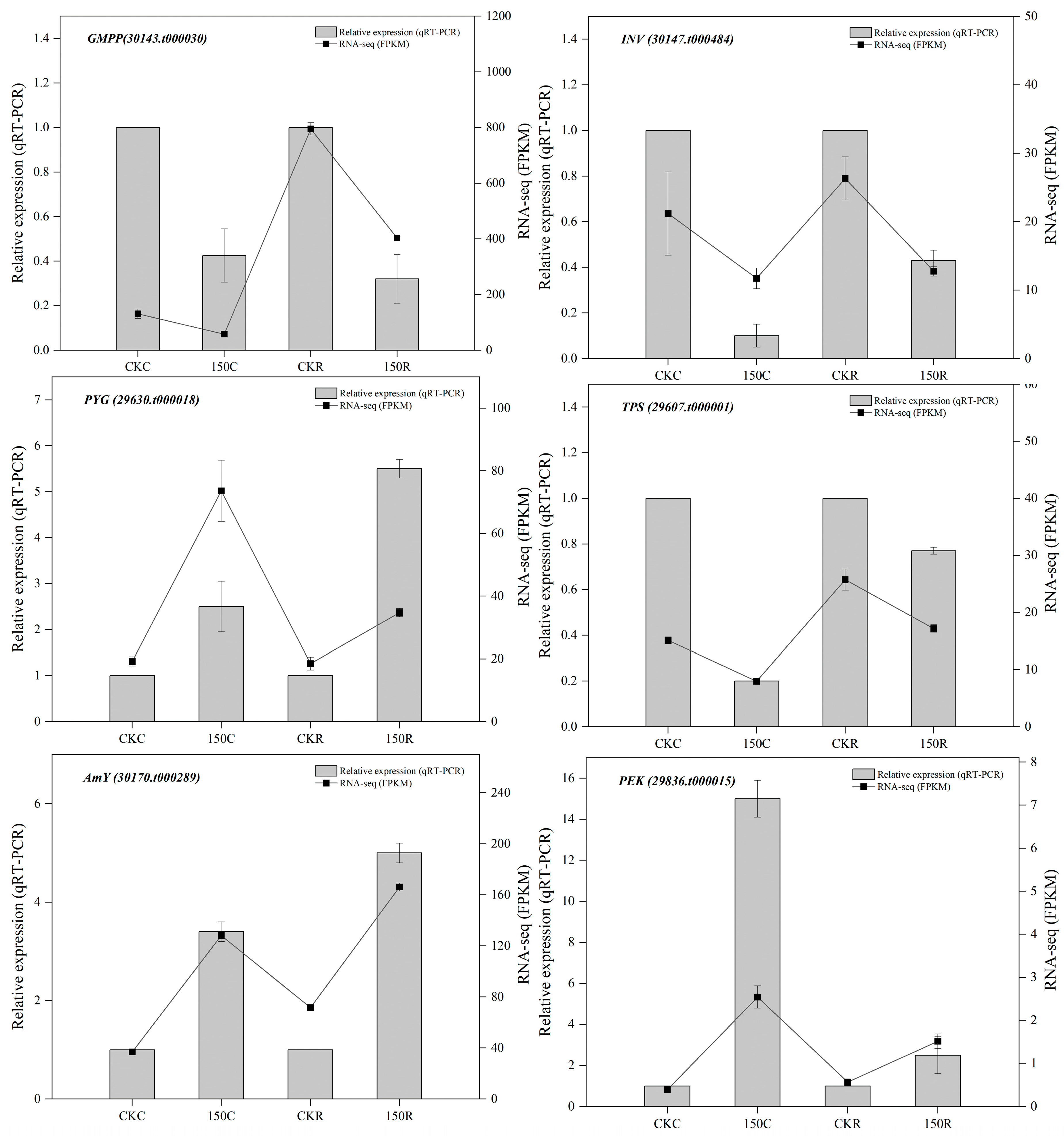
3.6. Validation of RNA-Seq by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Munns, R.; Gilliham, M. Salinity tolerance of crops–what is the cost? New Phytol. 2015, 208, 668–673. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.; Tu, W.; Mu, L.; Sun, Z.; Hu, Q.; Yang, Y. Saline alkali water desalination project in Southern Xinjiang of China: A review of desalination planning, desalination schemes and economic analysis. Renew. Sustain. Energy Rev. 2019, 113, 109268. [Google Scholar] [CrossRef]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guo, Y. Elucidating the molecular mechanisms mediating plant salt-stress responses. New Phytol. 2018, 217, 523–539. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innov. 2020, 1, 100017. [Google Scholar] [CrossRef] [PubMed]
- Mijiti, M.; Zhang, Y.; Zhang, C.; Wang, Y. Physiological and molecular responses of Betula platyphylla Suk to salt stress. Trees 2017, 31, 1653–1665. [Google Scholar] [CrossRef]
- Mo, S.; Biao, A.; Wang, Z.; Lin, S.; Yang, T.; Pan, L.; Zeng, S. Spatio transcriptome uncover novel insight into the Lycium ruthenicum seedling tolerant to salt stress. Ind. Crops Prod. 2022, 177, 114502. [Google Scholar] [CrossRef]
- Zhang, D.; He, S.; Fu, Y.; Yu, R.; Gao, X.; Wang, Z.; Chen, M. Transcriptome analysis reveals key genes in response to salinity stress during seed germination in Setaria italica. Environ. Exp. Bot. 2021, 191, 104604. [Google Scholar] [CrossRef]
- Li, H.; Lv, Q.; Deng, J.; Huang, J.; Cai, F.; Liang, C.; Chen, Q. Transcriptome analysis reveals key seed-development genes in common buckwheat (Fagopyrum esculentum). Int. J. Mol. Sci. 2019, 20, 4303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, P.L.; Li, S.X.; Yao, K.S.; Geng, H.Y.; Liu, J.Y.; Wang, Y.N.; Lin, J.X. Integrated metabolomic and transcriptomic strategies to reveal adaptive mechanisms in castor plant during germination stage under alkali stress. Environ. Exp. Bot. 2022, 203, 105031. [Google Scholar] [CrossRef]
- Hanley, M.E. Seedling herbivory, community composition and plant life history traits. Perspect. Plant Ecol. 1998, 1, 191–205. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, L.; Cao, X.; Liu, L.; Jiang, B.; Zhang, C.; Han, T. Cotyledons facilitate the adaptation of early-maturing soybean varieties to high-latitude long-day environments. Plant. Cell Environ. 2021, 44, 2551–2564. [Google Scholar] [CrossRef]
- Shi, S.; An, L.; Mao, J.; Aluko, O.O.; Ullah, Z.; Xu, F. The CBL interacting protein kinase NtCIPK23 positively regulates seed germination and early seedling development in tobacco (Nicotiana tabacum L.). Plants 2021, 10, 323. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, Y.; Peng, X.; Yan, J.; Yan, X.; Zhou, Z.; Lin, J. Cotyledon removal decreases salt tolerance during seedling establishment of Ricinus communis, an oilseed energy crop species. Ind. Crops Prod. 2019, 142, 111857. [Google Scholar] [CrossRef]
- Zhou, J.C.; Zhao, W.W.; Yin, C.H.; Song, J.; Wang, B.S.; Fan, J.L.; Feng, G. The role of cotyledons in the establishment of Suaeda physophora seedlings. Plant Biosyst. -Int. J. Deal. All Asp. Plant Biol. 2014, 148, 584–590. [Google Scholar]
- Wang, Y.N.; Peng, X.Y.; Hua, X.Y.; Zang, W.; Yang, X.F.; Lin, J.X. Research progress on the physiological function of plant cotyledons. Pratacultural. Sci. 2018, 35, 2988–2997. [Google Scholar]
- Ribeiro, P.R.; Fernandez, L.G.; Castro, R.D.; Ligterink, W.; Hilhorst, H.W. Physiological and biochemical responses of Ricinus communis seedlings to different temperatures: A metabolomics approach. BMC Plant Biol 2014, 14, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Lu, Y. Research advance in preparation and application of castor seed oil and its derivatives. China Oils Fats 2011, 36, 52–56. [Google Scholar]
- Baldwin, B.S.; Cossar, R.D. Castor yield in response to planting date at four locations in the south-central United States. Ind. Crops Prod. 2009, 29, 316–319. [Google Scholar] [CrossRef]
- El-Nashar, Y.I.; Hassan, B.A.; Aboelsaadat, E.M. Response of Nemesia (Nemesia×hybridus) plants to different irrigation water sources and arbuscular mycorrhizal fungi inoculation. Agric. Water Manag. 2021, 243, 106416. [Google Scholar] [CrossRef]
- Sarula, L.P. Research progress on adaptation of castor (Ricinus communis L.) to Abiotic Stress. J. Inn. Mong. Univ. Natl. 2021, 36, 135–139. [Google Scholar]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Han, R.C.; Liu, L.L.; Liu, J.L.; Xie, D.M.; Peng, D.Y.; Yu, N.J. Cloning and quantitative expression analysis of GMPP gene from Dendrobium huoshanense. China J. Chin. Mater. Med. 2019, 44, 1552–1557. [Google Scholar]
- Tie, Y.Y.; Tian, J. Cloning of sativum inwertase gene AsINV and its expression analysis in response to low temperature and drought stress. Plant Physiol. J. 2021, 57, 2258–2270. [Google Scholar]
- Zhang, Y.Y.; Lv, Y.; Xuan, W.Y.; Zhang, X.Q.; Xu, N.J.; Sun, X. Response of trehalose—6-phosphate synthetase (TPS) to abiotic stress in Gracilariopsis lemaneiformis. Oceanol. Et Limnol. Sin. 2021, 52, 777–785. [Google Scholar]
- Yue, C.; Cao, H.L.; Lin, H.Z.; Hu, J.; Ye, L.J.; Li, J.M.; Hao, Z.L.; Hao, X.Y.; Sun, Y.; Yang, Y.J.; et al. Expression patterns of alpha-amylase and beta-amylase genes provide insights into the molecular mechanisms underlying the responses of tea plants (Camellia sinensis) to stress and postharvest processing treatments. Planta 2019, 250, 281–298. [Google Scholar] [CrossRef]
- Wang, C.; Zhang, J.; Xie, Y. Cloning and hpRNAi Vector Construction of Brassica napus Pyrophosphate-dependent Phosphofructokinase (PFK). Mol. Plant Breed. 2006, 14, 199–204. [Google Scholar]
- Huang, F.L.; Jiang, Y.X.; Zhang, S.B.; Liu, S.; Eh, T.J.; Meng, F.J.; Lei, P. A Comparative Analysis on Morphological and Physiological Characteristics between Castor Varieties (Ricinus communis L.) under Salt Stress. Sustainability 2022, 14, 10032. [Google Scholar] [CrossRef]
- Diao, H.Y.; Wang, A.Z.; Yuan, F.H.; Guan, D.X.; Sun, Y.; Wu, J.B. Applications of compound-specific isotope analysis in tree non-structural carbohydrates research: A review. Ying Yong Sheng Tai Xue Bao= J. Appl. Ecol. 2020, 31, 4291–4300. [Google Scholar]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Tang, X.F.; Wu, Y.M.; Huang, Y.B.; Tang, Y.X. The roles of MYB transcription factors on plant defense responses and its molecular mechanism. Yi Chuan/Hered. 2008, 30, 1265–1271. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.; Liu, Z. Basic Helix-Loop-Helix (bHLH) transcription factor family in Yellow horn (Xanthoceras sorbifolia Bunge): Genome-wide characterization, chromosome location, phylogeny, structures and expression patterns. Int. J. Biol. Macromol. 2020, 160, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Liu, Z.; Hu, Y.; Kim, H.C.; Liu, S.; Liu, J.Q.; Xu, L.P.; Li, J.X.; Zhao, Y.; Yu, Z.L.; et al. Transcriptome analysis of salt stress responsiveness in the seedlings of wild and cultivated Ricinus communis L. J. Biotechnol. 2021, 327, 106–116. [Google Scholar] [CrossRef]
- Dai, X.; Xu, Y.; Ma, Q.; Xu, W.; Wang, T.; Xue, Y.; Chong, K. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol. 2007, 143, 1739–1751. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.; Tang, M.; Zhang, H.; Luo, D.; Cao, S.; Hu, Y.; Chen, P. The transcription factor HcERF4 confers salt and drought tolerance in kenaf (Hibiscus cannabinus L.). Plant Cell 2022, 150, 207–221. [Google Scholar] [CrossRef]
- Wei, J.; Liu, D.; Liu, Y.; Wei, S. Physiological analysis and transcriptome sequencing reveal the effects of salt stress on banana (Musa acuminata cv. BD) Leaf. Front. Plant Sci. 2022, 13, 822838. [Google Scholar] [CrossRef]
- Zhu, T.; Lin, J.; Zhang, M.; Li, L.; Zhao, C.; Chen, M. Phytohormone involved in salt tolerance regulation of Elaeagnus angustifolia L. seedlings. J. For. Res. 2019, 24, 235–242. [Google Scholar] [CrossRef]
- Zou, L.; Li, T.; Li, B.; He, J.; Liao, C.; Wang, L.; Wu, Q. De novo transcriptome analysis provides insights into the salt tolerance of Podocarpus macrophyllus under salinity stress. BMC Plant Biol. 2021, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Sicilia, A.; Testa, G.; Santoro, D.F.; Cosentino, S.L.; Lo Piero, A.R. RNA Seq analysis of giant cane reveals the leaf transcriptome dynamics under long-term salt stress. BMC Plant Biol. 2019, 19, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñiz García, M.N.; Cortelezzi, J.I.; Fumagalli, M.; Capiati, D.A. Expression of the Arabidopsis ABF4 gene in potato increases tuber yield, improves tuber quality and enhances salt and drought tolerance. Plant Mol. Biol. 2018, 98, 137–152. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Liu, J.N.; Yan, L.; Liang, Q.; Fang, H.; Wang, C.; Wu, D. Comparative transcriptome analysis unravels defense pathways of Fraxinus velutina Torr against salt stress. Front. Plant Sci. 2022, 13, 842726. [Google Scholar] [CrossRef]
- Moons, A.; Prinsen, E.; Bauw, G.; Van Montagu, M. Antagonistic effects of abscisic acid and jasmonates on salt stress-inducible transcripts in rice roots. Plant Cell 1997, 9, 2243–2259. [Google Scholar]
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Su, M.; Li, X.F.; Ma, X.Y.; Peng, X.J.; Zhao, A.G.; Cheng, L.Q.; Liu, G.S. Cloning two P5CS genes from bioenergy sorghum and their expression profiles under abiotic stresses and MeJA treatment. Plant Sci. 2011, 181, 652–659. [Google Scholar] [CrossRef]
- Iskandar, H.M.; Widyaningrum, D.; Suhandono, S. Cloning and characterization of P5CS1 and P5CS2 genes from Saccharum officinarum L. under drought stress. J. Trop. Crop Sci. 2014, 1, 23–30. [Google Scholar] [CrossRef]
- Geng, G.; Lv, C.; Stevanato, P.; Li, R.; Liu, H.; Yu, L.; Wang, Y. Transcriptome analysis of salt-sensitive and tolerant genotypes reveals salt-tolerance metabolic pathways in sugar beet. Int. J. Mol. Sci. 2019, 20, 5910. [Google Scholar] [CrossRef] [Green Version]
- Montero-Barrientos, M.; Hermosa, R.; Cardoza, R.E.; Gutierrez, S.; Nicolas, C.; Monte, E. Transgenic expression of the Trichoderma harzianum hsp70 gene increases Arabidopsis resistance to heat and other abiotic stresses. J. Plant Physiol. 2010, 167, 659–665. [Google Scholar] [CrossRef]
- Shafi, A.; Chauhan, R.; Gill, T.; Swarnkar, M.K.; Sreenivasulu, Y.; Kumar, S.; Singh, A.K. Expression of SOD and APX genes positively regulates secondary cell wall biosynthesis and promotes plant growth and yield in Arabidopsis under salt stress. Plant Mol. Biol. 2015, 87, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Jin, Y.; Li, M.; Dong, L.; Guo, D.; Lu, C.; Qi, H. Analysis of CmCADs and three lignifying enzymes in oriental melon (‘CaiHong7’) seedlings in response to three abiotic stresses. Sci. Hortic. 2018, 237, 257–268. [Google Scholar] [CrossRef]
- Sui, J.; Deng, H.; Yang, J.; Sun, J.; Qu, C. Cloning and expression characteristics of TsCAD1 gene under abiotic stress condition in Toona sinensis. Genom. Appl. Biol. 2019, 38, 4617–4625. [Google Scholar]
- Zhao, W.; Ding, L.; Liu, J.; Zhang, X.; Li, S.; Zhao, K.; Chen, F. Regulation of lignin biosynthesis by an atypical bHLH protein CmHLB in Chrysanthemum. J. Exp. Bot. 2022, 73, 2403–2419. [Google Scholar] [CrossRef]
- Guo, G.Y.; Bai, F.; Liu, W.; Mi, C.L. Advances in research of the regulation of transcription factors of lignin biosynthesis. Sci. Agric. Sin. 2015, 48, 1277–1287. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Forward Primer | Reverse Primer |
|---|---|---|
| AMY (30170.t000289) | CTTCAAGCTGCTGTGCAAGG | GTTCCTTCAGCCCCCAATCA |
| PFK (29836.t000015) | GGTATTGGCCTGGTGAAGCT | AACAACAAGAACCGCATGGC |
| PYG (29630.T000018) | GCACTGGTTCGCATTTGTGT | AAGACTCCGGTTACTTGGCG |
| TPS (29607.t000001) | GGGTGATGTTGTTTGGTGCC | CACTGACAAAATGCCGTGCA |
| GMPP (30143.t000030) | GGATGGACATTGGACAGCCA | ACACAGCCTGGACCTATTGC |
| INV (30147.t000484) | AGGTCCATTTGGGCTTCTGG | CGCAGCTTTTCATCGTCGAG |
| Sample | Raw Reads | Raw Bases | Clean Reads | Clean Bases | Valid Bases | Q30 | GC | Mapped Reads |
|---|---|---|---|---|---|---|---|---|
| CKC1 | 50.78M | 7.62G | 50.31M | 7.32G | 96.14% | 95.45% | 43.26% | 95.61% |
| CKC2 | 50.19M | 7.53G | 49.77M | 7.21G | 95.72% | 95.78% | 43.33% | 96.32% |
| CKC3 | 48.08M | 7.21G | 47.66M | 6.89G | 95.54% | 95.74% | 43.51% | 96.52% |
| CKR1 | 47.67M | 7.15G | 47.32M | 6.83G | 95.47% | 96.15% | 43.19% | 95.60% |
| CKR2 | 51.55M | 7.73G | 51.16M | 7.36G | 95.25% | 96.02% | 43.18% | 96.25% |
| CKR3 | 48.77M | 7.31G | 48.38M | 6.94G | 94.87% | 96.11% | 43.18% | 95.27% |
| S150C1 | 48.04M | 7.21G | 47.60M | 6.85G | 95.03% | 95.86% | 43.25% | 95.39% |
| S150C2 | 50.49M | 7.57G | 50.06M | 7.24G | 95.58% | 95.96% | 43.28% | 95.28% |
| S150C3 | 48.91M | 7.34G | 48.55M | 7.04G | 96.00% | 95.83% | 43.48% | 95.52% |
| S150R1 | 50.95M | 7.64G | 50.55M | 7.31G | 95.63% | 95.69% | 43.29% | 95.58% |
| S150R2 | 49.55M | 7.43G | 49.18M | 7.13G | 95.89% | 95.78% | 43.36% | 95.16% |
| S150R3 | 50.56M | 7.58G | 50.10M | 7.31G | 96.40% | 95.17% | 43.43% | 94.71% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, X.; Ma, Y.; Cheng, S.; Jin, Z.; Shi, C.; Liu, J.; Lin, J.; Yan, X. Castor Plant Adaptation to Salinity Stress during Early Seedling Stage by Physiological and Transcriptomic Methods. Agronomy 2023, 13, 693. https://doi.org/10.3390/agronomy13030693
Deng X, Ma Y, Cheng S, Jin Z, Shi C, Liu J, Lin J, Yan X. Castor Plant Adaptation to Salinity Stress during Early Seedling Stage by Physiological and Transcriptomic Methods. Agronomy. 2023; 13(3):693. https://doi.org/10.3390/agronomy13030693
Chicago/Turabian StyleDeng, Xiaoxia, Yuwen Ma, Shuang Cheng, Zixuan Jin, Congcong Shi, Junyu Liu, Jixiang Lin, and Xiufeng Yan. 2023. "Castor Plant Adaptation to Salinity Stress during Early Seedling Stage by Physiological and Transcriptomic Methods" Agronomy 13, no. 3: 693. https://doi.org/10.3390/agronomy13030693