
In Vitro Infection Dynamics of Japanese Encephalitis Virus in Established Porcine Cell Lines

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus and Cell Lines
2.2. Virus Growth Kinetics and Cell Viability
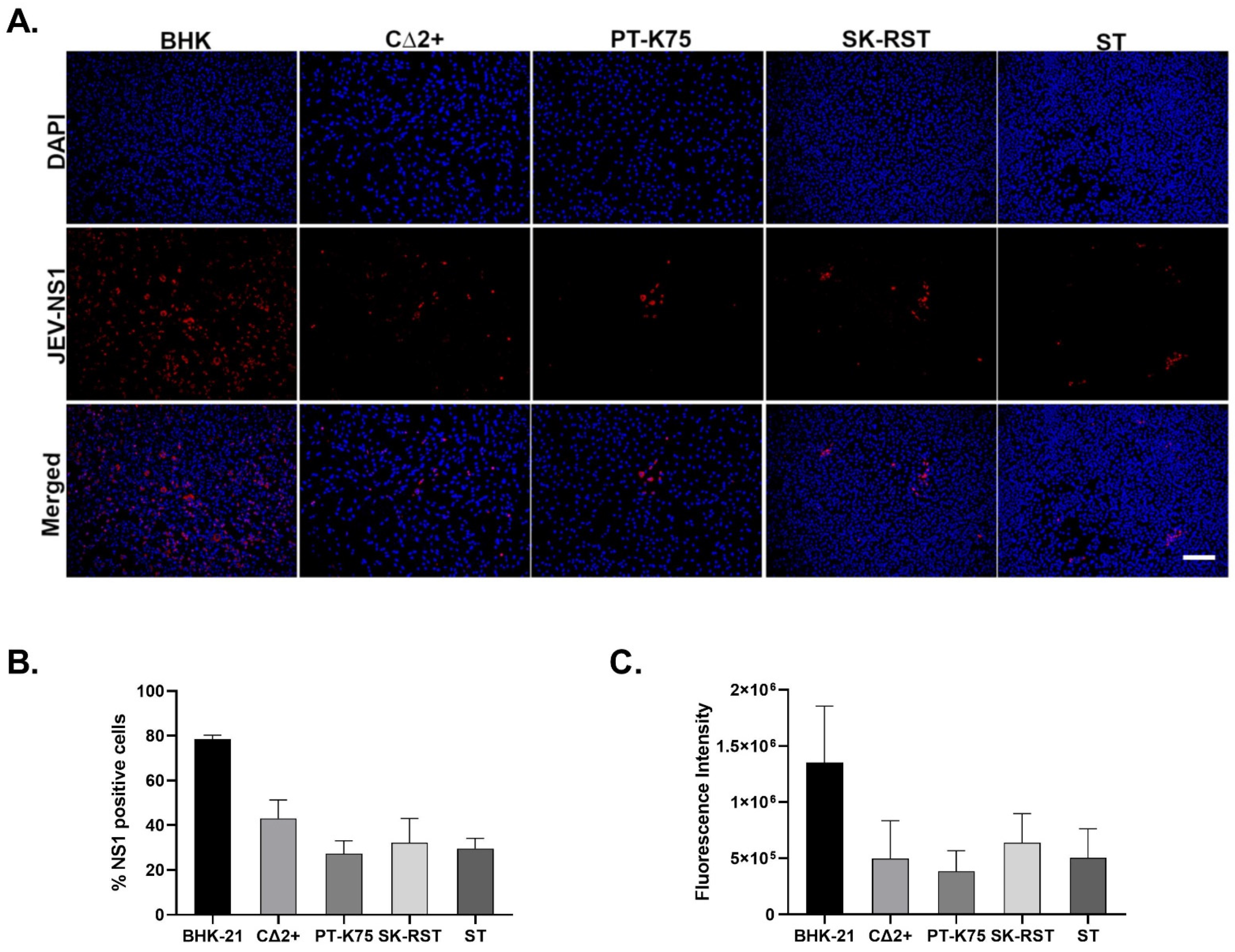
2.3. Immunofluorescence Assay
3. Results
3.1. Porcine Cell Lines Support JEV SA14-14-2 Replication and Produce Infectious Virus
3.2. JEV SA14-14-2 Induces Differential Cytopathic Changes in Porcine Cell Lines
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
References
- Daep, C.A.; Muñoz-Jordán, J.L.; Eugenin, E.A. Flaviviruses, an expanding threat in public health: Focus on dengue, West Nile, and Japanese encephalitis virus. J. Neurovirol. 2014, 20, 539–560. [Google Scholar] [CrossRef] [Green Version]
- Woolhouse, M.; Brierley, L. Epidemiological characteristics of human-infective RNA viruses. Sci. Data 2018, 5, 180017. [Google Scholar] [CrossRef] [PubMed]
- Campbell, G.L.; Hills, S.L.; Fischer, M.; Jacobson, J.A.; Hoke, C.H.; Hombach, J.M.; Marfin, A.A.; Solomon, T.; Tsai, T.F.; Tsu, V.D.; et al. Estimated global incidence of Japanese encephalitis. Bull. World Health Organ. 2011, 89, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Mathers, C.D.; Ezzati, M.; Lopez, A.D. Measuring the Burden of Neglected Tropical Diseases: The Global Burden of Disease Framework. PLoS Neglected Trop. Dis. 2007, 1, e114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turtle, L.; Solomon, T. Japanese encephalitis—The prospects for new treatments. Nat. Rev. Neurol. 2018, 14, 298–313. [Google Scholar] [CrossRef]
- Chai, C.; Wang, Q.; Cao, S.; Zhao, Q.; Wen, Y.; Huang, X.; Wen, X.; Yan, Q.; Ma, X.; Wu, R. Serological and molecular epidemiology of Japanese encephalitis virus infections in swine herds in China, 2006–2012. J. Veter. Sci. 2018, 19, 151–155. [Google Scholar] [CrossRef]
- De Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens Are Competent Vectors for Japanese Encephalitis Virus. PLoS Negl. Trop. Dis. 2017, 11, e0005294. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Faye, O.; Prot, M.; Casademont, I.; Fall, G.; Garcia, M.D.F.; Diagne, M.M.; Kipela, J.-M.; Fall, I.S.; Holmes, E.; et al. Autochthonous Japanese Encephalitis with Yellow Fever Coinfection in Africa. N. Engl. J. Med. 2017, 376, 1483–1485. [Google Scholar] [CrossRef]
- Preziuso, S.; Mari, S.; Mariotti, F.; Rossi, G. Detection of Japanese Encephalitis Virus in bone marrow of healthy young wild birds collected in 1997–2000 in Central Italy. Zoonoses Public Health 2018, 65, 798–804. [Google Scholar] [CrossRef]
- Liang, G.-D.; Huanyu, W. Epidemiology of Japanese encephalitis: Past, present, and future prospects. Ther. Clin. Risk Manag. 2015, 11, 435–448. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.-R.; Tesh, R.B.; Rico-Hesse, R. Genetic Variation of Japanese Encephalitis Virus in Nature. J. Gen. Virol. 1990, 71, 2915–2922. [Google Scholar] [CrossRef]
- Chen, W.-R.; Rico-Hesse, R.; Tesh, R.B. A new Genotype of Japanese Encephalitis Virus from Indonesia. Am. J. Trop. Med. Hyg. 1992, 47, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.S.; Cohnstaedt, L.W.; Noronha, L.E.; Mitzel, D.; McVey, D.S.; Cernicchiaro, N. Perspectives Regarding the Risk of Introduction of the Japanese Encephalitis Virus (JEV) in the United States. Front. Vet. Sci. 2020, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sharma, P.; Shukla, K.K.; Misra, S.; Nyati, K.K. Japanese encephalitis virus: Associated immune response and recent progress in vaccine development. Microb. Pathog. 2019, 136, 103678. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.-I.; Lee, Y.-M. Japanese encephalitis. Hum. Vaccines Immunother. 2013, 10, 263–279. [Google Scholar] [CrossRef] [Green Version]
- Quan, T.M.; Thao, T.T.N.; Duy, N.M.; Nhat, T.M.; Clapham, H. Estimates of the global burden of Japanese encephalitis and the impact of vaccination from 2000–2015. eLife 2020, 9, e51027. [Google Scholar] [CrossRef]
- Mansfield, K.L.; Hernández-Triana, L.M.; Banyard, A.C.; Fooks, A.R.; Johnson, N. Japanese encephalitis virus infection, diagnosis and control in domestic animals. Veter.-Microbiol. 2017, 201, 85–92. [Google Scholar] [CrossRef]
- Hurk, A.F.V.D.; Ritchie, S.A.; Mackenzie, J.S. Ecology and Geographical Expansion of Japanese Encephalitis Virus. Annu. Rev. Èntomol. 2009, 54, 17–35. [Google Scholar] [CrossRef] [Green Version]
- Pearce, J.C.; Learoyd, T.P.; Langendorf, B.J.; Logan, J.G. Japanese encephalitis: The vectors, ecology and potential for expansion. J. Travel Med. 2018, 25, S16–S26. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.-Y.; Bi, P.; Cazelles, B.; Zhou, S.; Huang, S.-Q.; Yang, J.; Pei, Y.; Wu, X.-X.; Fu, S.-H.; Tong, S.-L.; et al. How environmental conditions impact mosquito ecology and Japanese encephalitis: An eco-epidemiological approach. Environ. Int. 2015, 79, 17–24. [Google Scholar] [CrossRef]
- Weaver, S.C.; Barrett, A.D.T. Transmission cycles, host range, evolution and emergence of arboviral disease. Nat. Rev. Microbiol. 2004, 2, 789–801. [Google Scholar] [CrossRef] [PubMed]
- Erlanger, T.E.; Weiss, S.; Keiser, J.; Utzinger, J.; Wiedenmayer, K. Past, Present, and Future of Japanese Encephalitis. Emerg. Infect. Dis. 2009, 15, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Filgueira, L.; Lannes, N. Review of Emerging Japanese Encephalitis Virus: New Aspects and Concepts about Entry into the Brain and Inter-Cellular Spreading. Pathogens 2019, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Lyons, A.C.; Huang, Y.-J.S.; Park, S.L.; Ayers, V.B.; Hettenbach, S.M.; Higgs, S.; McVey, D.S.; Noronha, L.; Hsu, W.-W.; Vanlandingham, D.L. Shedding of Japanese Encephalitis Virus in Oral Fluid of Infected Swine. Vector-Borne Zoonotic Dis. 2018, 18, 469–474. [Google Scholar] [CrossRef]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar] [CrossRef] [Green Version]
- Ladreyt, H.; Durand, B.; Dussart, P.; Chevalier, V. How Central Is the Domestic Pig in the Epidemiological Cycle of Japanese Encephalitis Virus? A Review of Scientific Evidence and Implications for Disease Control. Viruses 2019, 11, 949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.L.; Huang, Y.-J.S.; Lyons, A.C.; Ayers, V.B.; Hettenbach, S.M.; McVey, D.S.; Burton, K.R.; Higgs, S.; VanLandingham, D.L. North American domestic pigs are susceptible to experimental infection with Japanese encephalitis virus. Sci. Rep. 2018, 8, 7951. [Google Scholar] [CrossRef]
- Ogasa, A.; Yokoki, Y.; Fujisaki, Y.; Habu, A. Reproductive disorders in boars infected experimentally with Japanese encephalitis virus. Japanese. J. Anim. Reprod. 1977, 23, 171–175. [Google Scholar] [CrossRef]
- Zheng, B.; Wang, X.; Liu, Y.; Li, Y.; Long, S.; Gu, C.; Ye, J.; Xie, S.; Cao, S. Japanese Encephalitis Virus infection induces inflammation of swine testis through RIG-I—NF-ĸB signaling pathway. Vet. Microbiol. 2019, 238, 108430. [Google Scholar] [CrossRef]
- Takashima, I.; Watanabe, T.; Ouchi, N.; Hashimoto, N. Ecological Studies of Japanese Encephalitis Virus in Hokkaido: Interepidemic Outbreaks of Swine Abortion and Evidence for the Virus to Overwinter Locally. Am. J. Trop. Med. Hyg. 1988, 38, 420–427. [Google Scholar] [CrossRef]
- Faizah, A.N.; Kobayashi, D.; Amoa-Bosompem, M.; Higa, Y.; Tsuda, Y.; Itokawa, K.; Miura, K.; Hirayama, K.; Sawabe, K.; Isawa, H. Evaluating the competence of the primary vector, Culex tritaeniorhynchus, and the invasive mosquito species, Aedes japonicus japonicus, in transmitting three Japanese encephalitis virus genotypes. PLoS Negl. Trop. Dis. 2020, 14, e0008986. [Google Scholar] [CrossRef] [PubMed]
- Boyer, S.; Peng, B.; Pang, S.; Chevalier, V.; Duong, V.; Gorman, C.; Dussart, P.; Fontenille, D.; Cappelle, J. Dynamics and diversity of mosquito vectors of Japanese encephalitis virus in Kandal province, Cambodia. J. Asia-Pac. Èntomol. 2020, 23, 1048–1054. [Google Scholar] [CrossRef]
- Oliveira, A.R.; Piaggio, J.; Cohnstaedt, L.W.; McVey, D.S.; Cernicchiaro, N. A quantitative risk assessment (QRA) of the risk of introduction of the Japanese encephalitis virus (JEV) in the United States via infected mosquitoes transported in aircraft and cargo ships. Prev. Vet. Med. 2018, 160, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.R.S.; Piaggio, J.; Cohnstaedt, L.W.; McVey, D.S.; Cernicchiaro, N. Introduction of the Japanese encephalitis virus ( JEV ) in the United States—A qualitative risk assessment. Transbound. Emerg. Dis. 2019, 66, 1558–1574. [Google Scholar] [CrossRef]
- Williams, D.T.; Lunt, R.A.; Wang, L.F.; Daniels, P.W.; Newberry, K.M.; MacKenzie, J.S. Experimental infections of pigs with Japanese encephalitis virus and closely related Australian flaviviruses. Am. J. Trop. Med. Hyg. 2001, 65, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Yamada, M.; Nakamura, K.; Yoshii, M.; Kaku, Y.; Narita, M. Brain Lesions Induced by Experimental Intranasal Infection of Japanese Encephalitis Virus in Piglets. J. Comp. Pathol. 2009, 141, 156–162. [Google Scholar] [CrossRef]
- Redant, V.; Favoreel, H.W.; Dallmeier, K.; Van Campe, W.; De Regge, N. Efficient control of Japanese encephalitis virus in the central nervous system of infected pigs occurs in the absence of a pronounced inflammatory immune response. J. Neuroinflamm. 2020, 17, 315. [Google Scholar] [CrossRef] [PubMed]
- Laureti, M.; Narayanan, D.; Rodriguez-Andres, J.; Fazakerley, J.K.; Kedzierski, L. Flavivirus Receptors: Diversity, Identity, and Cell Entry. Front. Immunol. 2018, 9, 2180. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Liu, H.; Xiao, T.; Wang, Z.; Nie, X.; Li, X.; Qian, P.; Qin, L.; Han, X.; Zhang, J.; et al. CRISPR screening of por-cine sgRNA library identifies host factors associated with Japanese encephalitis virus replication. Nat. Comm. 2020, 11, 5178. [Google Scholar] [CrossRef]
- Mukherjee, S.; Sengupta, N.; Chaudhuri, A.; Akbar, I.; Singh, N.; Chakraborty, S.; Suryawanshi, A.R.; Bhattacharyya, A.; Basu, A. PLVAP and GKN3 are two critical host cell receptors which facilitate Japanese encephalitis virus entry into neurons. Sci. Rep. 2018, 8, 11784. [Google Scholar] [CrossRef] [Green Version]
- García-Nicolás, O.; Braun, R.O.; Milona, P.; Lewandowska, M.; Dijkman, R.; Alves, M.P.; Summerfield, A. Targeting of the Nasal Mucosa by Japanese Encephalitis Virus for Non-Vector-Borne Transmission. J. Virol. 2018, 92, e01091-18. [Google Scholar] [CrossRef] [Green Version]
- García-Nicolás, O.; Lewandowska, M.; Ricklin, M.E.; Summerfield, A. Monocyte-Derived Dendritic Cells as Model to Evaluate Species Tropism of Mosquito-Borne Flaviviruses. Front. Cell. Infect. Microbiol. 2019, 9, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dulac, G.C.; Afshar, A. Porcine circovirus antigens in PK-15 cell line (ATCC CCL-33) and evidence of antibodies to circovirus in Canadian pigs. Can. J. Vet. Res. 1989, 53, 431–433. [Google Scholar] [PubMed]
- Shimizu, Y.; Furuuchi, S.; Hayashi, S.; Kumagai, T.; Sasahara, J. Porcine Kidney Cell Line Persistently Contaminated with Avirulent Swine Fever Virus. J. Gen. Virol. 1969, 4, 625–628. [Google Scholar] [CrossRef]
- Chitko-McKown, C.G.; Chapes, S.K.; Miller, L.C.; Riggs, P.K.; Ortega, M.T.; Green, B.T.; McKown, R.D. Development and characterization of two porcine monocyte-derived macrophage cell lines. Results Immunol. 2013, 3, 26–32. [Google Scholar] [CrossRef] [Green Version]
- Fadnis, P.R.; Ravi, V.; Desai, A.; Turtle, L.; Solomon, T. Innate Immune Mechanisms in Japanese Encephalitis Virus Infection: Effect on Transcription of Pattern Recognition Receptors in Mouse Neuronal Cells and Brain Tissue. Viral Immunol. 2013, 26, 366–377. [Google Scholar] [CrossRef] [Green Version]
- Takamatsu, Y.; Okamoto, K.; Dinh, D.T.; Yu, F.; Hayasaka, D.; Uchida, L.; Nabeshima, T.; Buerano, C.C.; Morita, K. NS1′ protein expression facilitates production of Japanese encephalitis virus in avian cells and embryonated chicken eggs. J. Gen. Virol. 2014, 95, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, Z.-R.; Zhang, Y.-N.; Liu, J.; Deng, C.-L.; Shi, P.-Y.; Yuan, Z.-M.; Ye, H.-Q.; Zhang, B. A replication-defective Japanese encephalitis virus (JEV) vaccine candidate with NS1 deletion confers dual protection against JEV and West Nile virus in mice. NPJ Vaccines 2020, 5, 73. [Google Scholar] [CrossRef]
- Shen, T.; Liu, K.; Miao, D.; Cao, R.; Chen, P. Effective inhibition of Japanese encephalitis virus replication by shRNAs targeting various viral genes in vitro and in vivo. Virology 2014, 454–455, 48–59. [Google Scholar] [CrossRef] [Green Version]
- Song, B.-H.; Yun, G.-N.; Kim, J.-K.; Yun, S.-I.; Lee, Y.-M. Biological and genetic properties of SA14-14-2, a live-attenuated Japanese encephalitis vaccine that is currently available for humans. J. Microbiol. 2012, 50, 698–706. [Google Scholar] [CrossRef]
- Yun, S.-I.; Song, B.-H.; Kim, J.-K.; Yun, G.-N.; Lee, E.-Y.; Li, L.; Kuhn, R.J.; Rossmann, M.G.; Morrey, J.D.; Lee, Y.-M. A Molecularly Cloned, Live-Attenuated Japanese Encephalitis Vaccine SA14-14-2 Virus: A Conserved Single Amino Acid in the ij Hairpin of the Viral E Glycoprotein Determines Neurovirulence in Mice. PLoS Pathog. 2014, 10, e1004290. [Google Scholar] [CrossRef] [Green Version]
- Khromykh, A.A.; Sedlak, P.L.; Guyatt, K.J.; Hall, R.A.; Westaway, E.G. Efficient trans -Complementation of the Flavivirus Kunjin NS5 Protein but Not of the NS1 Protein Requires Its Coexpression with Other Components of the Viral Replicase. J. Virol. 1999, 73, 10272–10280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindenbach, B.D.; Rice, C.M. trans-Complementation of yellow fever virus NS1 reveals a role in early RNA replication. J. Virol. 1997, 71, 9608–9617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Xu, A.; Wu, X.; Zhang, Y.; Guo, Y.; Guo, F.; Pan, Z.; Kong, L. Japanese encephalitis virus induces apoptosis by the IRE1/JNK pathway of ER stress response in BHK-21 cells. Arch. Virol. 2016, 161, 699–703. [Google Scholar] [CrossRef]
- Ueba, N.; Kimura, T.; Kimoto, T. Multinucleated Giant Cell Formation in BHK-21-528 Cell Monolayers Infected with Japanese Encephalitis Viruses 1. Jpn. J. Microbiol. 1976, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gromowski, G.D.; Firestone, C.-Y.; Whitehead, S.S. Genetic Determinants of Japanese Encephalitis Virus Vaccine Strain SA14-14-2 That Govern Attenuation of Virulence in Mice. J. Virol. 2015, 89, 6328–6337. [Google Scholar] [CrossRef] [Green Version]
- Yun, S.-I.; Song, B.-H.; Polejaeva, I.; Davies, C.J.; White, K.L.; Lee, Y.-M. Comparison of the live-attenuated Japanese encephalitis vaccine SA14 -14-2 strain with its pre-attenuated virulent parent SA14 strain: Similarities and differences in vitro and in vivo. J. Gen. Virol. 2016, 97, 2575–2591. [Google Scholar] [CrossRef]
- German, A.C.; Myint, K.S.A.; Mai, N.T.H.; Pomeroy, I.; Phu, N.H.; Tzartos, J.; Winter, P.; Collett, J.; Farrar, J.; Barrett, A.; et al. A preliminary neuropathological study of Japanese encephalitis in humans and a mouse model. Trans. R. Soc. Trop. Med. Hyg. 2006, 100, 1135–1145. [Google Scholar] [CrossRef]
- Lannes, N.; Summerfield, A.; Filgueira, L. Regulation of inflammation in Japanese encephalitis. J. Neuroinflamm. 2017, 14, 1–11. [Google Scholar] [CrossRef]
- Hsieh, J.T.; John, A.L.S. Japanese encephalitis virus and its mechanisms of neuroinvasion. PLoS Pathog. 2020, 16, e1008260. [Google Scholar] [CrossRef]
- Sooryanarain, H.; Sapkal, G.N.; Gore, M.M. Pathogenic and vaccine strains of Japanese encephalitis virus elicit different levels of human macrophage effector functions. Arch. Virol. 2012, 157, 1905–1918. [Google Scholar] [CrossRef] [PubMed]
- Donne, E.; Pasmans, F.; Boyen, F.; Van Immerseel, F.; Adriaensen, C.; Hernalsteens, J.; Ducatelle, R.; Haesebrouck, F. Survival of Salmonella serovar Typhimurium inside porcine monocytes is associated with complement binding and suppression of the production of reactive oxygen species. Vet. Microbiol. 2005, 107, 205–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maria-Pilar, J.D.B.; Dudal, S.; Dornand, J.; Gross, A. Cellular bioterrorism: How Brucella corrupts macrophage physiology to promote invasion and proliferation. Clin. Immunol. 2005, 114, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Van Reeth, K.; Adair, B. Macrophages and respiratory viruses. Pathol. Biol. 1997, 45, 184–192. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Peak Titer ± SD (pfu/mL) | Time of Peak Titer (hpi) |
|---|---|---|
| BHK-21 | 4.83 ± 2.4 × 107 | 48 |
| C∆2+ | 4.11 ± 5.54 × 106 | 48 |
| SK-RST | 5.83 ± 1.44 × 106 | 48 |
| PT-K75 | 6.38 ± 8.08 × 104 | 72 |
| ST | 5.22 ± 3.22 × 106 | 120 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adetunji, S.A.; Smolensky, D.; Mitzel, D.N.; Owens, J.L.; Chitko-McKown, C.G.; Cernicchiaro, N.; Noronha, L.E. In Vitro Infection Dynamics of Japanese Encephalitis Virus in Established Porcine Cell Lines. Pathogens 2021, 10, 1468. https://doi.org/10.3390/pathogens10111468
Adetunji SA, Smolensky D, Mitzel DN, Owens JL, Chitko-McKown CG, Cernicchiaro N, Noronha LE. In Vitro Infection Dynamics of Japanese Encephalitis Virus in Established Porcine Cell Lines. Pathogens. 2021; 10(11):1468. https://doi.org/10.3390/pathogens10111468
Chicago/Turabian StyleAdetunji, Shakirat A., Dmitriy Smolensky, Dana N. Mitzel, Jeana L. Owens, Carol G. Chitko-McKown, Natalia Cernicchiaro, and Leela E. Noronha. 2021. "In Vitro Infection Dynamics of Japanese Encephalitis Virus in Established Porcine Cell Lines" Pathogens 10, no. 11: 1468. https://doi.org/10.3390/pathogens10111468