
Prevalence of Borrelia burgdorferi Sensu Lato in Ixodes ricinus Ticks Collected from Kaylaka Park in Pleven, Bulgaria

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Tick Collection
2.3. Geographic Integration
2.4. DNA Extraction and PCR
2.5. Borrelia burgdorferi s.l. DNA Detection by Polymerase Chain Reaction (PCR)
2.6. Agarose Gel Electrophoresis and Imaging
2.7. Dark-Field Microscopy (DFM)
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Beaujean, D.; Crutzen, R.; Kengen, C.; van Steenbergen, J.; Ruwaard, D. Increase in Ticks and Lyme Borreliosis, Yet Research into Its Prevention on the Wane. Vector-Borne Zoonotic Dis. 2016, 16, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Vandekerckhove, O.; De Buck, E.; Van Wijngaerden, E. Lyme disease in Western Europe: An emerging problem? A systematic review. Acta Clin. Belg. 2021, 76, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Sigal, L. Lyme Borreliosis. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Stupica, D.; Stanek, G.; Strle, F. Lyme Borreliosis. In Emerging Infectious Diseases; Ergönül, Ö., Can, F., Madoff, L., Akova, M., Eds.; Academic Press: Amsterdam, The Netherlands, 2014; pp. 323–335. [Google Scholar]
- Stone, B.L.; Tourand, Y.; Brissette, C.A. Brave New Worlds: The Expanding Universe of Lyme Disease. Vector-Borne Zoonotic Dis. 2017, 17, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Sykes, R.A.; Makiello, P. An estimate of Lyme borreliosis incidence in Western Europe. J. Public Health 2016, 39, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junttila, J.; Peltomaa, M.; Soini, H.; Marjamäki, M.; Viljanen, M.K. Prevalence of Borrelia burgdorferi in Ixodes ricinus Ticks in Urban Recreational Areas of Helsinki. J. Clin. Microbiol. 1999, 37, 1361–1365. [Google Scholar] [CrossRef] [Green Version]
- Christova, I.; Dimitrov, H.; Trifonova, I.; Gladnishka, T.; Mitkovska, V.; Stojanova, A.; Taseva, E.; Ivanova, V. Detection of Human Tick-borne Pathogens in Rodents from Bulgaria. Acta Zool. Bulg. 2012, 4, 111–114. [Google Scholar]
- Christova, I.; Schouls, L.; van de Pol, I.; Park, J.; Panayotov, S.; Lefterova, V.; Kantardjiev, T.; Dumler, J.S. High Prevalence of Granulocytic Ehrlichiae and Borrelia burgdorferi Sensu Lato in Ixodes ricinus Ticks from Bulgaria. J. Clin. Microbiol. 2001, 39, 4172–4174. [Google Scholar] [CrossRef] [Green Version]
- Sormunen, J.J.; Kulha, N.; Klemola, T.; Mäkelä, S.; Vesilahti, E.; Vesterinen, E.J. Enhanced threat of tick-borne infections within cities? Assessing public health risks due to ticks in urban green spaces in Helsinki, Finland. Zoonoses Public Health 2020, 67, 823–839. [Google Scholar] [CrossRef]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M.; et al. Driving forces for changes in geographical distribution of Ixodes ricinus ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Rauter, C.; Hartung, T. Prevalence of Borrelia burgdorferi Sensu Lato Genospecies in Ixodes ricinus Ticks in Europe: A Metaanalysis. Appl. Environ. Microbiol. 2005, 71, 7203–7216. [Google Scholar] [CrossRef] [Green Version]
- Blazhev, A.; Karcheva, M.; Cenova, A.; Blazheva, S.; Kostov, K. Serological Tests for Lyme Borreliosis. Gen. Med. 2018, 20, 20–24. [Google Scholar]
- Hansford, K.M.; Fonville, M.; Gillingham, E.L.; Coipan, E.C.; Pietzsch, M.E.; Krawczyk, A.I.; Vaux, A.G.; Cull, B.; Sprong, H.; Medlock, J.M. Ticks and Borrelia in urban and peri-urban green space habitats in a city in southern England. Ticks Tick-Borne Dis. 2017, 8, 353–361. [Google Scholar] [CrossRef] [PubMed]
- Klemola, T.; Sormunen, J.; Mojzer, J.; Mäkelä, S.; Vesterinen, E.J. High tick abundance and diversity of tick-borne pathogens in a Finnish city. Urban Ecosyst. 2019, 22, 817–826. [Google Scholar] [CrossRef] [Green Version]
- Kowalec, M.; Szewczyk, T.; Welc-Falęciak, R.; Siński, E.; Karbowiak, G.; Bajer, A. Ticks and the city—Are there any differences between city parks and natural forests in terms of tick abundance and prevalence of spirochaetes? Parasites Vectors 2017, 10, 573. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, C.P.; Heutschi, D.; Lenz, N.; Tischhauser, W.; Péter, O.; Rais, O.; Beuret, C.M.; Leib, S.L.; Bankoul, S.; Ackermann-Gäumann, R. Prevalence of tick-borne pathogens in questing Ixodes ricinus ticks in urban and suburban areas of Switzerland. Parasites Vectors 2017, 10, 558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schorn, S.; Pfister, K.; Reulen, H.; Mahling, M.; Silaghi, C. Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasites Vectors 2011, 4, 135. [Google Scholar] [CrossRef] [Green Version]
- Tomanović, S.; Radulović, Ž.; Masuzawa, T.; Milutinović, M. Coexistence of emerging bacterial pathogens in Ixodes ricinus ticks in Serbia. Parasite 2010, 17, 211–217. [Google Scholar] [CrossRef]
- Žákovská, A.; Vostal, K.; Martiníková, H. A longitudinal study of the prevalence of borreliae in ticks in the urban locality of Brno—Pisárky, Czech Republic. J. Vector Ecol. 2008, 33, 385–388. [Google Scholar] [CrossRef]
- Žákovská, A.; Nejezchlebová, H.; Bartoňková, N.; Rašovská, T.; Kučerová, H.; Norek, A.; Ovesná, P. Activity of the tick Ixodes Ricinus monitored in a suburban park in Brno, Czech Republic, in association with the evaluation of selected repellents. J. Vector Ecol. 2013, 38, 295–300. [Google Scholar] [CrossRef]
- Christova, I.; Van De Pol, J.; Yazar, S.; Velo, E.; Schouls, L. Identification of Borrelia burgdorferi sensu lato, Anaplasma and Ehrlichia Species, and Spotted Fever Group Rickettsiae in Ticks from Southeastern Europe. Eur. J. Clin. Microbiol. 2003, 22, 535–542. [Google Scholar] [CrossRef]
- Blazhev, A.; Atanasova, M.; Kostov, K.; Doychinova, T.; Blazheva, S.; Karcheva, M. Estimation of Ixodes ricinus (Acari: Ixodidae) Populations of Kaylaka Park in the Town of Pleven, Bulgaria. Insects 2021, 12, 808. [Google Scholar] [CrossRef] [PubMed]
- Karcheva, M.; Blazhev, A.; Tsenova, A.; Atanasova, M.; Kostov, K. Retrospective seroepidemiological survey of Lyme borreliosis in Pleven region, Bulgaria. Eur. J. Public Health 2018, 28, cky218.125. [Google Scholar] [CrossRef]
- Angelov, V. Association “Hunting and Fishing Society—Pleven” From Its Foundation to the Present Day, 1st ed.; Borisova, G., Ed.; University Press St. Kliment Ohridski: Sofia, Bulgaria, 2019; p. 340. [Google Scholar]
- Georgieva, G.; Gecheva, G. Fauna Bulgaria 32; Beron, P., Ed.; Marin Drinov Academic Publishing House: Sofia, Bulgaria, 2013; Volume 32, p. 226. [Google Scholar]
- Estrada-Peña, A.; Mihalca, A.D.; Petney, T.N. Ticks of Europe and North Africa: A Guide to Species Identification; Springer: Berlin/Heidelberg, Germany, 2018. [Google Scholar] [CrossRef]
- Chu, C.-Y.; Jiang, B.-G.; Liu, W.; Zhao, Q.-M.; Wu, X.-M.; Zhang, P.-H.; Zhan, L.; Yang, H.; Cao, W.-C. Presence of pathogenic Borrelia burgdorferisensu lato in ticks and rodents in Zhejiang, south-east China. J. Med. Microbiol. 2008, 57, 980–985. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.; Gazumyan, A.; Schwartz, I. rRNA gene organization in the Lyme disease spirochete, Borrelia burgdorferi. J. Bacteriol. 1992, 174, 3757–3765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Postic, D.; Assous, M.V.; Grimont, P.A.D.; Baranton, G. Diversity of Borrelia burgdorfeii Sensu Lato Evidenced by Restriction Fragment Length Polymorphism of rrf (5S)-rrl (23S) Intergenic Spacer Amplicons. Int. J. Syst. Bacteriol. 1994, 44, 743–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wills, M.K.B.; Kirby, A.M.; Lloyd, V.K. Detecting the Lyme Disease Spirochete, Borrelia Burgdorferi, in Ticks Using Nested PCR. J. Vis. Exp. 2018, 132, e56471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chvostáč, M.; Špitalská, E.; Václav, R.; Vaculová, T.; Minichová, L.; Derdáková, M. Seasonal Patterns in the Prevalence and Diversity of Tick-Borne Borrelia burgdorferi Sensu Lato, Anaplasma phagocytophilum and Rickettsia spp. in an Urban Temperate Forest in South Western Slovakia. Int. J. Environ. Res. Public Health 2018, 15, 994. [Google Scholar] [CrossRef] [Green Version]
- Hodžić, A.; Fuehrer, H.-P.; Duscher, G.G. First Molecular Evidence of Zoonotic Bacteria in Ticks in Bosnia and Herzegovina. Transbound. Emerg. Dis. 2016, 64, 1313–1316. [Google Scholar] [CrossRef]
- Coipan, E.C.; Vladimirescu, A.F. Ixodes ricinus ticks (Acari: Ixodidae): Vectors for Lyme disease spirochetes in Romania. Exp. Appl. Acarol. 2011, 54, 293–300. [Google Scholar] [CrossRef]
- Kalmár, Z.; Mihalca, A.D.; Dumitrache, M.O.; Gherman, C.M.; Magdaş, C.; Mircean, V.; Oltean, M.; Domşa, C.; Matei, I.A.; Mărcuţan, D.I.; et al. Geographical distribution and prevalence of Borrelia burgdorferi genospecies in questing Ixodes ricinus from Romania: A countrywide study. Ticks Tick-borne Dis. 2013, 4, 403–408. [Google Scholar] [CrossRef]
- Sen, E.; Uchishima, Y.; Okamoto, Y.; Fukui, T.; Kadosaka, T.; Ohashi, N.; Masuzawa, T. Molecular detection of Anaplasma phagocytophilum and Borrelia burgdorferi in Ixodes ricinus ticks from Istanbul metropolitan area and rural Trakya (Thrace) region of north-western Turkey. Ticks Tick-borne Dis. 2011, 2, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Milutinović, M.; Masuzawa, T.; Tomanović, S.; Radulović, Ž.; Fukui, T.; Okamoto, Y. Borrelia burgdorferi sensu lato, Anaplasma phagocytophilum, Francisella tularensis and their co-infections in host-seeking Ixodes ricinus ticks collected in Serbia. Exp. Appl. Acarol. 2008, 45, 171–183. [Google Scholar] [CrossRef] [PubMed]
- Strnad, M.; Honig, V.; Růžek, D.; Grubhoffer, L.; Rego, R. Europe-Wide Meta-Analysis of Borrelia burgdorferi Sensu Lato Prevalence in Questing Ixodes ricinus Ticks. Appl. Environ. Microbiol. 2017, 83, e00609-17. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.S.; Dautel, H.; Peña, A.E.; Kahl, O.; Lindgren, E. Effects of Climate Change on Ticks and Tick-Borne Diseases in Europe. Interdiscip. Perspect. Infect. Dis. 2009, 2009, 593232. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, D.; Gebhardt, R.; Petney, T.; Norra, S. On the complexity of measuring forests microclimate and interpreting its relevance in habitat ecology: The example of Ixodes ricinus ticks. Parasites Vectors 2017, 10, 549. [Google Scholar] [CrossRef] [Green Version]
- Hubálek, Z.; Halouzka, J. Prevalence rates of Borrelia burgdorferi sensu lato in host-seeking Ixodes ricinus ticks in Europe. Parasitol. Res. 1998, 84, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, K.; Szczotko, M.; Dmitryjuk, M. Borrelia miyamotoi—An Emerging Human Tick-Borne Pathogen in Europe. Microorganisms 2021, 9, 154. [Google Scholar] [CrossRef]
- Aenishaenslin, C.; Ravel, A.; Michel, P.; Gern, L.; Milord, F.; Waaub, J.-P.; Bélanger, D. From Lyme disease emergence to endemicity: A cross sectional comparative study of risk perceptions in different populations. BMC Public Health 2014, 14, 1–13. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Primer Name | Gene | Target Sequence (5′-3′) | Amplicon Size | Annealing Temperature | References |
|---|---|---|---|---|---|
| Primer 23S3 Out | rrf (5S)-rrl (23S) intergenic spacer | CGACCTTCTTCGCCTTAAAGC | 411 bp | 55 °C | Chu et al., 2008 [28] Schwartz et al., 1992 [29] |
| Primer 23Sa Out | TAAGCTGACTAATACTAATTA CCC | ||||
| Primer 1 In | rrf (5S)-rrl (23S) intergenic spacer | CTG CGA GTT CGC GGG AGA | 254 bp | 59 °C | Postic et al., 1994 [30] |
| Primer 2 In | TCC TAG GCA TTC ACC ATA | ||||
| FlaB Out Fw | FlaB | GCATCACTTTCAGGGTCTCA | 503 bp | 55 °C | Wills et al. [31] |
| FlaB Out Rv | TGGGGAACTTGATTAGCCTG | ||||
| FlaB In Fw | CTTTAAGAGTTCATGTTGGAG | 447 bp | 58 °C | ||
| FlaB In Rv | TCATTGCCATTGCAGATTGT |
| Year | Stage_Gender | Total | |||
|---|---|---|---|---|---|
| Male | Female | Nymph | Larva | ||
| 2017 | 58 | 99 | 12 | 0 | 169 |
| 2018 | 71 | 51 | 10 | 1 | 133 |
| 2019 | 109 | 103 | 32 | 0 | 244 |
| Total | 238 | 253 | 54 | 1 | 546 |
| Area | Type of Test | Detection of Borrelia sp. in I. ricinus by Any Test Percentage of Tick Infection (%) and Number of Positive Samples (n) | |||
|---|---|---|---|---|---|
| Number of Collected Ticks | PCR 23S/5S | PCR FlaB | DFM | ||
| Tick Infection (%) and Number of Positive to Tested Samples (n) | Percentage of Tick Infection (%) and Number of Positive to Tested Samples (n) | Percentage of Tick Infection (%) and Number of Positive to Tested Samples (n) | |||
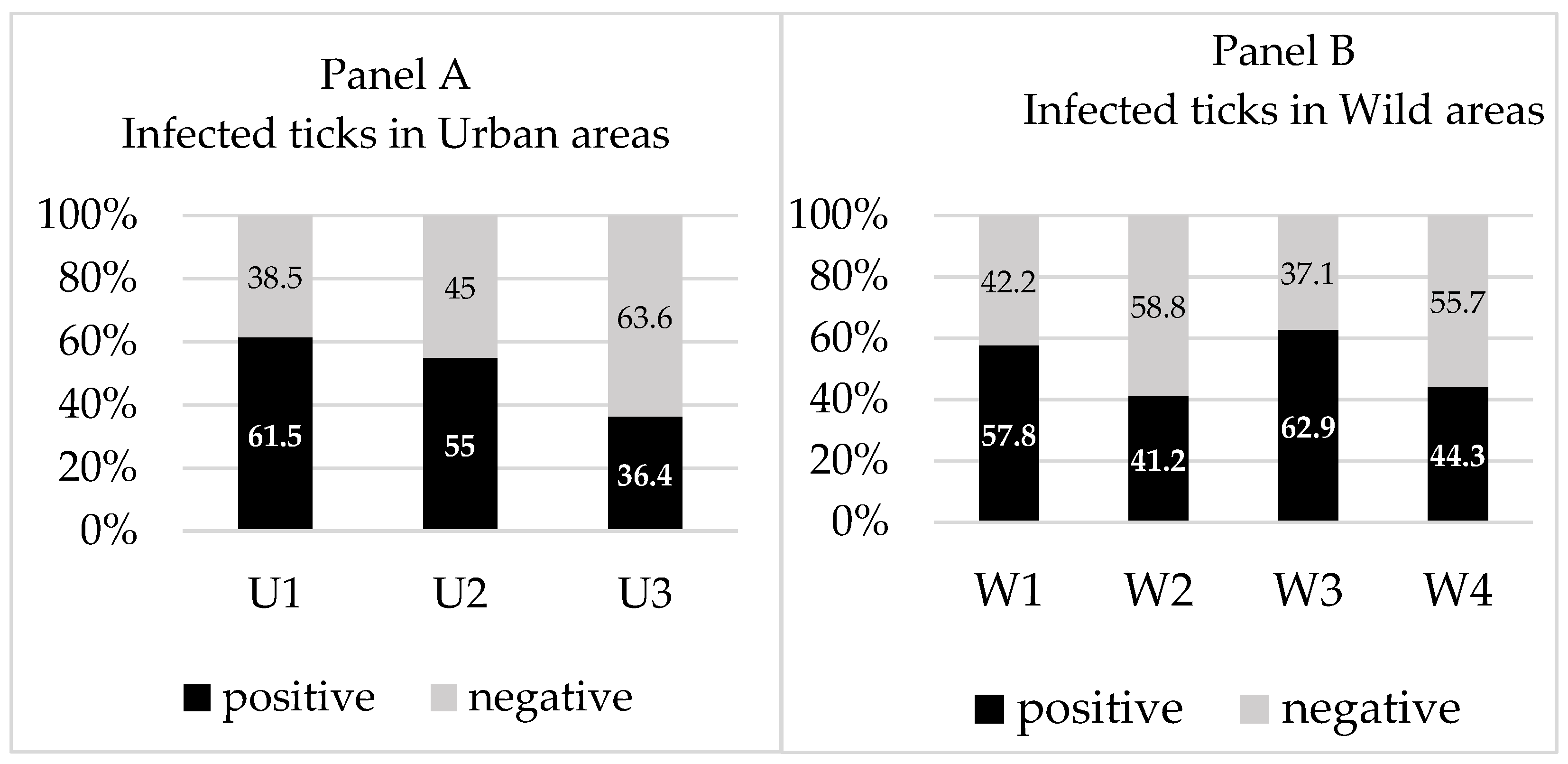
| Urban | 192 | 35.7 (n = 15/42) | 47.6 (n = 20/42) | 42 (n = 22/52) | 44.3 (n = 39) |
| U1 | 40 | 45.5 (n = 5/11) | 54.5 (n = 6/11) | 100 (n = 2/2) | 61.5 (n = 8) |
| U2 | 50 | 0 (n = 0/3) | 33.3 (n = 1/3) | 58.8 (n = 10/17) | 55.0 (n = 11) |
| U3 | 102 | 35.7 (n = 10/28) | 46.4 (n = 13/28) | 30.3 (n = 10/33) | 36.4 (n = 20) |
| Wild | 354 | 45 (n = 50/111) | 49.5 (n = 55/111) | 37.4 (n = 64/163) | 48.8 (n = 120) |
| W1 | 79 | 53.7 (n = 22/41) | 53.7 (n = 22/41) | 25 (n = 1/4) | 57.8 (n = 26) |
| W2 | 58 | 16.7 (n = 2/12) | 33.3 (n = 4/12) | 40.9 (n = 18/44) | 41.2 (n = 21) |
| W3 | 83 | 52.9 (n = 9/17) | 58.8 (n = 10/17) | 66.7 (n = 12/18) | 62.9 (n = 22) |
| W4 | 134 | 41.5 (n = 17/41) | 46.3 (n = 19/41) | 34 (n = 33/97) | 44.3 (n = 51) |
| Total | 546 | 42.5 (n = 65/153) | 49 (n = 75/153) | 40 (n = 86/218) | 47.6 (n = 159) |
| Year of Collection | Total | Method | Tested Ticks | Results | |
|---|---|---|---|---|---|
| Collected Ticks | Positive | Negative | |||
| 2017 | 169 | DFM | 124 | 49 (39.5%) | 75 (60.5%) |
| nPCR5S-23Sigs | 16 | 5 (31.3%) | 11 (68.8%) | ||
| nPCRFlaB | 16 | 8 (50.0%) | 8 (50.0%) | ||
| 2018 | 133 | DFM | 77 | 33 (42.9%) | 44 (57.1%) |
| nPCR5S-23Sigs | 28 | 8 (28.6%) | 20 (71.4%) | ||
| nPCRFlaB | 28 | 10 (35.7%) | 18 (64.3%) | ||
| 2019 | 244 | DFM | 14 | 4 (28.6%) | 10 (71.4%) |
| nPCR5S-23Sigs | 109 | 52 (47.7%) | 57 (52.3%) | ||
| nPCRFlaB | 109 | 57 (52.3%) | 52 (47.7%) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blazhev, A.; Stanilov, I.; Miteva, L.D.; Atanasova, M.; Blazheva, S.; Stanilova, S. Prevalence of Borrelia burgdorferi Sensu Lato in Ixodes ricinus Ticks Collected from Kaylaka Park in Pleven, Bulgaria. Microorganisms 2022, 10, 772. https://doi.org/10.3390/microorganisms10040772
Blazhev A, Stanilov I, Miteva LD, Atanasova M, Blazheva S, Stanilova S. Prevalence of Borrelia burgdorferi Sensu Lato in Ixodes ricinus Ticks Collected from Kaylaka Park in Pleven, Bulgaria. Microorganisms. 2022; 10(4):772. https://doi.org/10.3390/microorganisms10040772
Chicago/Turabian StyleBlazhev, Alexander, Iskren Stanilov, Lyuba Dineva Miteva, Milena Atanasova, Svetla Blazheva, and Spaska Stanilova. 2022. "Prevalence of Borrelia burgdorferi Sensu Lato in Ixodes ricinus Ticks Collected from Kaylaka Park in Pleven, Bulgaria" Microorganisms 10, no. 4: 772. https://doi.org/10.3390/microorganisms10040772