
Quinoa in Ecuador: Recent Advances under Global Expansion

 ,
,  , , ,
, , ,  ,
,
Abstract
:1. Introduction
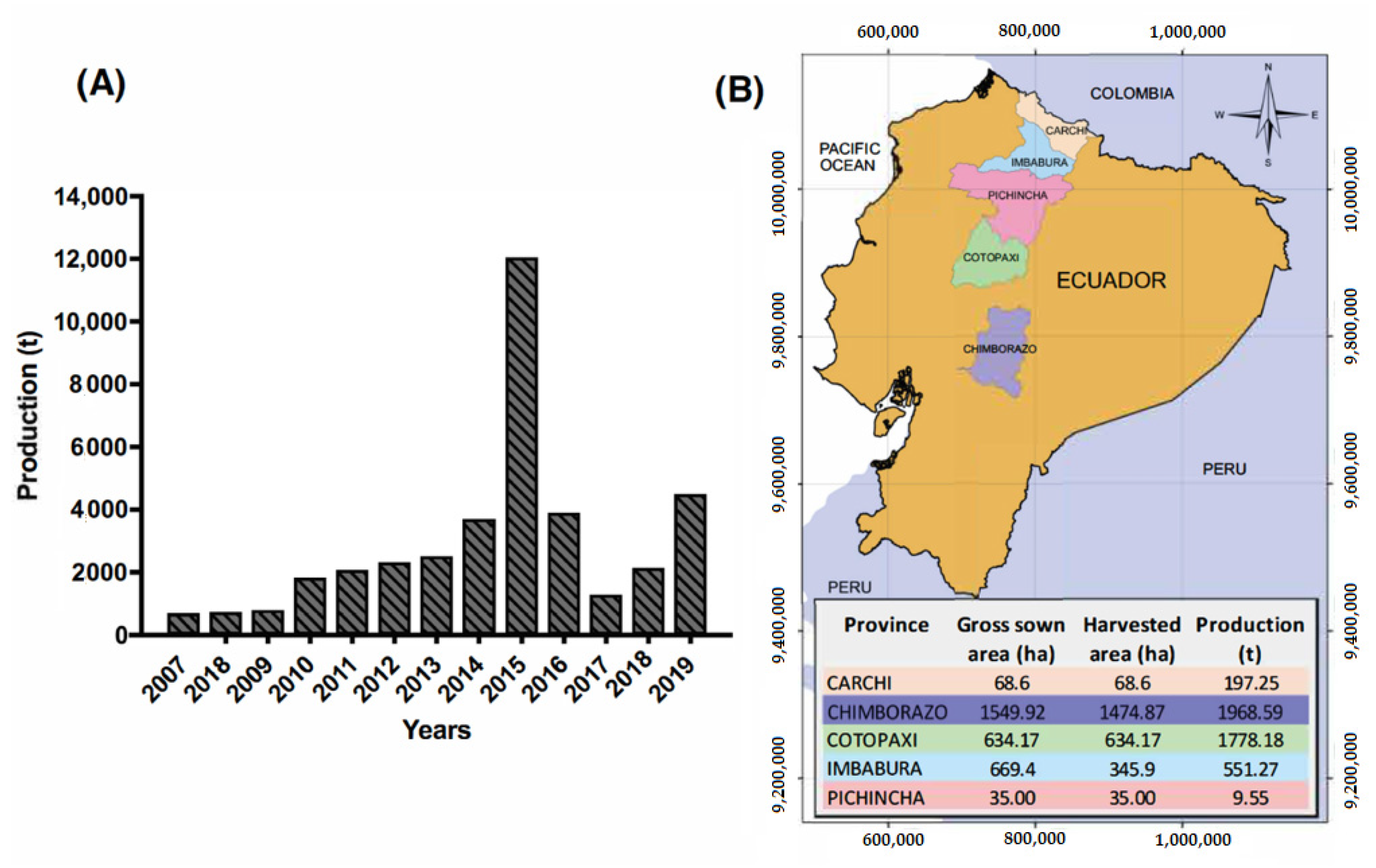
2. Quinoa Production in Ecuador
2.1. Quinoa Breeding in Ecuador
2.2. Participatory Research
2.3. Traditional Seed Production
2.4. The Nongovernmental Organization (NGO) “European Committee for Training and Agriculture” (CEFA) in Chimborazo
2.4.1. Quinoa Chain in Chimborazo
2.4.2. Main Actions of the PCV-IS in the Province of Chimborazo
Creation of a Technical Committee of Quinoa in Chimborazo
Promotion of Participatory Research and Links with the academy to Improve the Dynamics of Innovation in the Value Chain
Identification and Validation of Three Species for Intercropping with Quinoa
Use of Local Organic Matter Source for Soil Improvement
Development and Implementation of Adaptation Measures to Climate Change
3. Pests and Diseases
3.1. Quinoa Pests in Ecuador
3.2. Quinoa Diseases in Ecuador
3.2.1. Downy Mildew
3.2.2. Minor Diseases
4. Genetic Diversity
Wild Quinoa Relatives
5. Quinoa Adaptation to Tropical Areas
6. Quinoa Processing and Agroindustry
Quinoa Transformation
7. Participatory Plant Breeding Project between WSU and APROSANAMY (Case Study)
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jarvis, D.E.; Ho, Y.S.; Lightfoot, D.J.; Schmöckel, S.M.; Li, B.; Borm, T.J.A.; Ohyanagi, H.; Mineta, K.; Michell, C.T.; Saber, N.; et al. The Genome of Chenopodium quinoa. Nature 2017, 542, 307–312. [Google Scholar] [CrossRef] [Green Version]
- Tapia, M. The Long Journey of Quinoa: Who Wrote Its History. In State of the Art Report on Quinoa Around the World 2013; Bazile, D., Bertero, H.D., Nieto, C., Eds.; FAO: Rome, Italy; CIRAD: Paris, France, 2015; Volume 1, pp. 1–7. ISBN 978-92-5-108558-5. [Google Scholar]
- Hinojosa, L.; González, J.A.; Barrios-Masias, F.H.; Fuentes, F.; Murphy, K.M. Quinoa Abiotic Stress Responses: A Review. Plants 2018, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Adolf, V.I.; Jacobsen, S.-E.; Shabala, S. Salt Tolerance Mechanisms in Quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 2013, 92, 43–54. [Google Scholar] [CrossRef]
- Ruiz, K.B.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.-E. Quinoa—A Model Crop for Understanding Salt-Tolerance Mechanisms in Halophytes. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Bosque Sanchez, H.; Lemeur, R.; van Damme, P.; Jacobsen, S.-E. Ecophysiological Analysis of Drought and Salinity Stress of Quinoa (Chenopodium quinoa Willd.). Food Rev. Int. 2003, 19, 111–119. [Google Scholar] [CrossRef]
- Hinojosa, L.; Matanguihan, J.B.; Murphy, K.M. Effect of High Temperature on Pollen Morphology, Plant Growth and Seed Yield in Quinoa (Chenopodium quinoa Willd.). J. Agron. Crop Sci. 2019, 205, 33–45. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa, L.; Kumar, N.; Gill, K.S.; Murphy, K.M. Spectral Reflectance Indices and Physiological Parameters in Quinoa under Contrasting Irrigation Regimes. Crop Sci. 2019, 59, 1927–1944. [Google Scholar] [CrossRef] [Green Version]
- Hinojosa, L.; Sanad, M.N.M.E.; Jarvis, D.E.; Steel, P.; Murphy, K.; Smertenko, A. Impact of Heat and Drought Stress on Peroxisome Proliferation in Quinoa. Plant J. 2019, 99, 1144–1158. [Google Scholar] [CrossRef] [PubMed]
- Yang, A.; Akhtar, S.S.; Amjad, M.; Iqbal, S.; Jacobsen, S.-E. Growth and Physiological Responses of Quinoa to Drought and Temperature Stress. J. Agron. Crop Sci. 2016, 202, 445–453. [Google Scholar] [CrossRef]
- Craine, E.B.; Murphy, K.M. Seed Composition and Amino Acid Profiles for Quinoa Grown in Washington State. Front. Nutr. 2020, 7, 26. [Google Scholar] [CrossRef]
- Wu, G. Nutritional Properties of Quinoa. In Quinoa: Improvement and Sustainable Production; Murphy, K., Matanguihan, J., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 193–210. ISBN 978-1-118-62804-1. [Google Scholar]
- Vilcacundo, R.; Hernández-Ledesma, B. Nutritional and Biological Value of Quinoa (Chenopodium quinoa Willd.). Curr. Opin. Food Sci. 2017, 14, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Noratto, G.D.; Murphy, K.; Chew, B.P. Quinoa Intake Reduces Plasma and Liver Cholesterol, Lessens Obesity-Associated Inflammation, and Helps to Prevent Hepatic Steatosis in Obese Db/Db Mouse. Food Chem. 2019, 287, 107–114. [Google Scholar] [CrossRef]
- Bazile, D.; Jacobsen, S.-E.; Verniau, A. The Global Expansion of Quinoa: Trends and Limits. Front. Plant Sci. 2016, 7, 622. [Google Scholar] [CrossRef] [Green Version]
- Alandia, G.; Rodriguez, J.P.; Jacobsen, S.-E.; Bazile, D.; Condori, B. Global Expansion of Quinoa and Challenges for the Andean Region. Glob. Food Secur. 2020, 26, 100429. [Google Scholar] [CrossRef]
- FAO. FAOSTAT: Food and Agriculture Data. Crop Statistics. Available online: http://www.fao.org/faostat (accessed on 14 August 2020).
- Penafiel, D.; Vanhove, W.; Espinel, R.L.; Van Damme, P. Food Biodiversity Includes Both Locally Cultivated and Wild Food Species in Guasaganda, Central Ecuador. J. Ethn. Foods 2019, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Mestanza-Ramón, C.; Henkanaththegedara, S.M.; Vásconez Duchicela, P.; Vargas Tierras, Y.; Sánchez Capa, M.; Constante Mejía, D.; Jimenez Gutierrez, M.; Charco Guamán, M.; Mestanza Ramón, P. In-Situ and Ex-Situ Biodiversity Conservation in Ecuador: A Review of Policies, Actions and Challenges. Diversity 2020, 12, 315. [Google Scholar] [CrossRef]
- Zurita-Silva, A.; Fuentes, F.; Zamora, P.; Jacobsen, S.-E.; Schwember, A.R. Breeding Quinoa (Chenopodium quinoa Willd.): Potential and Perspectives. Mol. Breed. 2014, 34, 13–30. [Google Scholar] [CrossRef]
- Peralta, I.E.; Mazon, N. Quinoa in Ecuador. In State of the Art Report of Quinoa in the World in 2013; Bazile, D., Bertero, D., Nieto, C., Eds.; FAO: Rome, Italy; CIRAD: Paris, France, 2015; ISBN 978-92-5-108558-5. [Google Scholar]
- McElhinny, E.; Peralta, E.; Mazón, N.; Danial, D.L.; Thiele, G.; Lindhout, P. Aspects of Participatory Plant Breeding for Quinoa in Marginal Areas of Ecuador. Euphytica 2007, 153, 373–384. [Google Scholar] [CrossRef]
- Mazón, N.; Peralta, E.; Murillo, Á.; Rivera, M.; Guzmán, A.; Pichazaca, N.; Nicklin, C. It’s Not Just the Technology, Its the Surrounding System: How Researchers in Ecuador Found Ways to Make Themselves Useful to Farmers through Quinoa and Lupin Seed Systems. Exp. Agric. 2019, 55, 107–124. [Google Scholar] [CrossRef] [Green Version]
- Basantes-Morales, E.R.; Alconada, M.M.; Pantoja, J.L. Quinoa (Chenopodium quinoa Willd) Production in the Andean Region: Challenges and Potentials. J. Exp. Agric. Int. 2019, 1–18. [Google Scholar] [CrossRef]
- INEC (Instituto Nacional de Estadísticas y Censos), ESPAC (Encuesta de Superficie y Producción Agropecuaria Continua). Available online: https://www.ecuadorencifras.gob.ec/estadisticas-agropecuarias-2 (accessed on 12 September 2020).
- Cárdenas Oleas, S.E. Case Study: A Sustainable Livelihoods Analysis of a Non-Governmental Organization Working with Quinoa Producers in the Province of Chimborazo, Ecuador. Master’s Thesis, Iowa State University, Ames, IA, USA, 2015; p. 7896981. [Google Scholar]
- Murillo, A.; Mazon, N.; Rodriguez, L.; Vega, L. Mejoramiento Genético de Quinua (Chenopodium quinoa Willd.) en Ecuador. In Proceedings of the 7th Congreso Mundial de la Quinua y Otros Granos Andinos, Santiago, Chile, 25–28 March 2019. [Google Scholar]
- Monar, C.; Silva, D. Informe Anual de Actividades. Programa de Investigación y Producción de Semillas; Universidad Estatal de Bolívar, Facultad de Ciencias Agropecuarias Recursos Naturales y Del Ambiente: Guaranda, Ecuador, 2014. [Google Scholar]
- Sistema Nacional de Información. Available online: https://sni.gob.ec/inicio (accessed on 16 November 2020).
- Informe de Rendimientos Objetivos Quinua. 2019. Available online: http://sipa.agricultura.gob.ec/index.php/informe-de-rendimientos-objetivos/rendimiento-de-quinua (accessed on 16 November 2020).
- Ruales, C.; Fiallo, E. Plagas. In Quinua: Hacia su Cultivo Comercial; Wahli, C., Ed.; Latinreco S.A: Quito, Ecuador, 1990; p. 71. ISBN 978-9978-9901-3-1. [Google Scholar]
- Jacobsen, S.-E. Cultivo de Granos Andinos en Ecuador: Informe Sobre Los Rubros Quinua, Chocho y Amaranto; Editorial Abya Yala: Quito, Ecuador, 2002; ISBN 978-9978-22-258-4. [Google Scholar]
- Ministerio de Agricultura y Ganadería del Ecuador. Programa Nacional de Protección Vegetal. In Inventario de Plagas, Enfermedades y Malezas Del Ecuador: Quinoa; Ministerio de Agricultura y Ganadería Quito: Quito, Ecuador, 1986; p. 152. [Google Scholar]
- Orellana, H.; Peralta, E.; Castillo, R. Enfermedades de Quinoa. In Proceedings of the Curso de Quinoa, Quito, Ecuador, 16–18 October 1985; pp. 1–5. [Google Scholar]
- Danielsen, S.; Bonifacio, A.; Ames, T. Diseases of Quinoa (Chenopodium quinoa). Food Rev. Int. 2003, 19, 43–59. [Google Scholar] [CrossRef]
- Choi, Y.-J.; Danielsen, S.; Lübeck, M.; Hong, S.-B.; Delhey, R.; Shin, H.-D. Morphological and Molecular Characterization of the Causal Agent of Downy Mildew on Quinoa (Chenopodium quinoa). Mycopathologia 2010, 169, 403–412. [Google Scholar] [CrossRef]
- Baiswar, P.; Chandra, S.; Kumar, R.; Ngachan, S.V. Peronospora variabilis on Chenopodium murale in India. Australas. Plant Dis. Notes 2010, 5, 45–47. [Google Scholar] [CrossRef] [Green Version]
- Danielsen, S. Heterothallism in Peronospora farinosa f.sp. chenopodii, the Causal Agent of Downy Mildew of Quinoa (Chenopodium quinoa). J. Basic Microbiol. 2001, 41, 305–309. [Google Scholar] [CrossRef]
- Danielsen, S.; Mercado, V.H.; Ames, T.; Munk, L. Seed Transmission of Downy Mildew (Peronospora farinosa f. sp. chenopodii) in Quinoa and Effect of Relative Humidity on Seedling Infection. Seed Sci. Technol. 2004, 32, 91–98. [Google Scholar] [CrossRef]
- Testen, A.L.; del Mar Jiménez-Gasco, M.; Ochoa, J.B.; Backman, P.A. Molecular Detection of Peronospora variabilis in Quinoa Seed and Phylogeny of the Quinoa Downy Mildew Pathogen in South America and the United States. Phytopathology 2013, 104, 379–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobs, T. Postulation of Virulence Groups and Resistance Factors in the Quinoa/Downy Mildew Pathosystem Using Material from Ecuador. Plant Pathol. 1999, 48, 425–430. [Google Scholar] [CrossRef]
- Johnson, R. A Critical Analysis of Durable Resistance. Annu. Rev. Phytopathol. 1984, 22, 309–330. [Google Scholar] [CrossRef]
- Norwood, J.M.; Crute, I.R.; Johnson, A.G.; Gordon, P.L. A Demonstration of the Inheritance of Field Resistance to Lettuce Downy Mildew (Bremia lactucae Regel.) in Progeny Derived from Cv. Grand Rapids. Euphytica 1983, 32, 161–170. [Google Scholar] [CrossRef]
- Grube, R.C.; Ochoa, O.E. Comparative Genetic Analysis of Field Resistance to Downy Mildew in the Lettuce Cultivars “Grand Rapids” and “Iceberg”. Euphytica 2005, 142, 205–215. [Google Scholar] [CrossRef]
- Parlevliet, J.E. What is Durable Resistance, A General Outline. In Durability of Disease Resistance. Current Plant Science and Biotechnology in Agriculture; Jacobs, T., Parlevliet, J.E., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 23–39. ISBN 978-94-011-2004-3. [Google Scholar]
- Testen, A.L.; McKemy, J.M.; Backman, P.A. First Report of Passalora Leaf Spot of Quinoa Caused by Passalora dubia in the United States. Plant Dis. 2012, 97, 139. [Google Scholar] [CrossRef] [PubMed]
- Bertero, H.D.; King, R.W.; Hall, A.J. Photoperiod-Sensitive Development Phases in Quinoa (Chenopodium quinoa Willd.). Field Crops Res. 1999, 60, 231–243. [Google Scholar] [CrossRef]
- Mazón, N.; Rivera, M.M.; Peralta, I.E.; Estrella Engelmann, J.; Tapia, B.C. Catálogo del Banco de Germoplasma de Quinua (Chenoodium Quinoa Willd.) de INIAP-Ecuador; INIAP: Quito, Ecuador, 2001. [Google Scholar]
- Delgado, A.; Tapia, B.C.; Naranjo, E.; Borja, E.; Martin Clemente, J.P. Variación de la Agrobiodiversidad en Variedades Tradicionales de Quinua (Chenopodium Quinoa Willd) de la Región Andina de Ecuador. In Proceedings of the 7th Congreso Mundial de la Quinua y otros Granos Andinos, Iquique, Chile, 25–28 March 2017; p. 41. [Google Scholar]
- Tapia, B.C.; Peralta, E.; Mazón, N. Colecciones Núcleo de Quinua (Chenopodium Quinoa Willd.) del Banco de Germoplasma del INIAP, Ecuador. Axioma 2014, 2, 5–9. [Google Scholar]
- Jarvis, D.E.; Kopp, O.R.; Jellen, E.N.; Mallory, M.A.; Pattee, J.; Bonifacio, A.; Coleman, C.E.; Stevens, M.R.; Fairbanks, D.J.; Maughan, P.J. Simple Sequence Repeat Marker Development and Genetic Mapping in Quinoa (Chenopodium quinoa Willd.). J. Genet. 2008, 87, 39–51. [Google Scholar] [CrossRef]
- Christensen, S.A.; Pratt, D.B.; Pratt, C.; Nelson, P.T.; Stevens, M.R.; Jellen, E.N.; Coleman, C.E.; Fairbanks, D.J.; Bonifacio, A.; Maughan, P.J. Assessment of Genetic Diversity in the USDA and CIP-FAO International Nursery Collections of Quinoa (Chenopodium quinoa Willd.) Using Microsatellite Markers. Plant Genet. Resour. 2007, 5, 82. [Google Scholar] [CrossRef] [Green Version]
- Salazar, J.; de Lourdes Torres, M.; Gutierrez, B.; Torres, A.F. Molecular Characterization of Ecuadorian Quinoa (Chenopodium quinoa Willd.) Diversity: Implications for Conservation and Breeding. Euphytica 2019, 215, 60. [Google Scholar] [CrossRef] [Green Version]
- Wilson, H.D. Quinua Biosystematics II: Free-Living Populations. Econ. Bot. 1988, 42, 478–494. [Google Scholar] [CrossRef]
- Jellen, E.N.; Kolano, B.A.; Sederberg, M.C.; Bonifacio, A.; Maughan, P.J. Chenopodium. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 35–61. ISBN 978-3-642-14386-1. [Google Scholar]
- Navarrete, S.; Suasnavas, B. Análisis Filogenético de Quinua Silvestres (Chenopodium Quinoa Willd.) Utilizando el Gen rpoB en Muestras Provenientes de las Provincias de Carchi, Imabura, Cotopaxi y Chimborazo; Biotecnología de los Recursos Naturales, Salesian Polytechnic University: Quito, Ecuador, 2015. [Google Scholar]
- Jellen, E.N.; Jarvis, D.E.; Hunt, S.P.; Mangelsen, H.H.; Maughan, P.J. New Seed Collections of North American Pitseed Goosefoot (Chenopodium berlandieri) and Efforts to Identify Its Diploid Ancestors through Whole-Genome Sequencing. Cienc. Investig. Agrar. 2019, 46, 187–196. [Google Scholar] [CrossRef]
- Yao, S.; Lan, H.; Zhang, F. Variation of Seed Heteromorphism in Chenopodium album and the Effect of Salinity Stress on the Descendants. Ann. Bot. 2010, 105, 1015–1025. [Google Scholar] [CrossRef]
- Wilson, H.; Manhart, J. Crop/Weed Gene Flow: Chenopodium quinoa Willd. and C. berlandieri Moq. Theor. Appl. Genet. 1993, 86, 642–648. [Google Scholar] [CrossRef]
- Murphy, K.M.; Matanguihan, J.B.; Fuentes, F.F.; Gómez-Pando, L.R.; Jellen, E.N.; Maughan, P.J.; Jarvis, D.E. Quinoa Breeding and Genomics. In Plant Breeding Reviews; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 257–320. ISBN 978-1-119-52135-8. [Google Scholar]
- Hernández Medina, P.; Hernández Maqueda, R.; Gavilanes, P.; Marin Quevedo, K.P. Culture and Healthy Lifestyle: Factors Influencing the Decision to Buy Quinoa in the City of Latacunga in Cotopaxi Province, Ecuador. J. Food Prod. Mark. 2020, 26, 440–455. [Google Scholar] [CrossRef]
- Peralta, E.; Murillo, A.; Mazón , N.; Rodriguez Ortega, D.G.; Minchala, L.; Dominguez Olmedo, D.E.; Mina Chala, D.F. Evaluación de 239 Accesiones y 30 Líneas de Quinua (Chenopodium Quinoa Willd.) en Las Condiciones Agroecológicas de la Península de Santa Elena, Ecuador. In Proceedings of the 5th Congreso Mundial de la Quinua, 2nd Simposio Internacional de Granos Andinos, San Salvador de Jujuy, Argentina, 27–30 May 2015; FAO: Rome, Italy; p. 13. [Google Scholar]
- Angeli, V.; Miguel Silva, P.; Crispim Massuela, D.; Khan, M.W.; Hamar, A.; Khajehei, F.; Graeff-Hönninger, S.; Piatti, C. Quinoa (Chenopodium quinoa Willd.): An Overview of the Potentials of the “Golden Grain” and Socio-Economic and Environmental Aspects of Its Cultivation and Marketization. Foods 2020, 9, 216. [Google Scholar] [CrossRef] [Green Version]
- Madrid, D.; Salgado, E.; Verdugo, G.; Olguín, P.; Bilalis, D.; Fuentes, F. Morphological Traits Defining Breeding Criteria for Coastal Quinoa in Chile. Not. Bot. Horti Agrobot. Cluj-Napoca 2018, 46, 190–196. [Google Scholar] [CrossRef] [Green Version]
- Apaza, V.; Cáceres, G.; Estrada, R.; Pinedo, R. Catalogue of Commercial Varieties of Quinoa in Peru: A Future Planted Thousands of Years a Ago; FAO: Rome, Italy, 2015; ISBN 978-92-5-108765-7. [Google Scholar]
- Mestanza Uquillas, C.A.; Zambrano Calderón, K.; Pinargote Alava, J.; Veliz Zamora, D.; Vásconez Montufar, G.; Fernández-García, N.; Olmos, E. Evaluación agronómica de genotipos de quinua (Chenopodium quinoa Willd.) En condiciones agroclimáticas en la zona de mocache. Cienc. Tecnol. 2019, 12, 19–30. [Google Scholar] [CrossRef]
- Mestanza Uquillas, C.; Santana Chávez, J.; Veliz Zamora, D.; Vasconez, G. Rendimiento de grano de genotipos de quinua (Chenopodium quinoa Wild) sembrado a chorro continua, en el campus “La Maria.”. Univ. Cienc. Tecnol. 2020, 1, 53–60. [Google Scholar]
- Tang, Y.; Li, X.; Chen, P.X.; Zhang, B.; Hernandez, M.; Zhang, H.; Marcone, M.F.; Liu, R.; Tsao, R. Lipids, Tocopherols, and Carotenoids in Leaves of Amaranth and Quinoa Cultivars and a New Approach to Overall Evaluation of Nutritional Quality Traits. J. Agric. Food Chem. 2014, 62, 12610–12619. [Google Scholar] [CrossRef]
- Schoenlechner, R.; Drausinger, J.; Ottenschlaeger, V.; Jurackova, K.; Berghofer, E. Functional Properties of Gluten-Free Pasta Produced from Amaranth, Quinoa and Buckwheat. Plant Foods Hum. Nutr. Dordr. Neth. 2010, 65, 339–349. [Google Scholar] [CrossRef]
- Altuna, J.L.; Silva, M.; Álvarez, M.; Quinteros, M.F.; Morales, D.; Carrillo, W. Ecuadorian Quinoa (Chenopodium quinoa Willd) Fatty Acids Profile. Asian J. Pharm. Clin. Res. 2018, 11, 209. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Miralles, B.; Carrillo, W.; Hernández-Ledesma, B. In Vitro Chemopreventive Properties of Peptides Released from Quinoa (Chenopodium quinoa Willd.) Protein under Simulated Gastrointestinal Digestion. Food Res. Int. 2018, 105, 403–411. [Google Scholar] [CrossRef]
- Vilcacundo, R.; Martínez-Villaluenga, C.; Hernández-Ledesma, B. Release of Dipeptidyl Peptidase IV, α-Amylase and α-Glucosidase Inhibitory Peptides from Quinoa (Chenopodium quinoa Willd.) during in Vitro Simulated Gastrointestinal Digestion. J. Funct. Foods 2017, 35, 531–539. [Google Scholar] [CrossRef] [Green Version]
- Fiallos-Jurado, J.; Pollier, J.; Moses, T.; Arendt, P.; Barriga-Medina, N.; Morillo, E.; Arahana, V.; de Lourdes Torres, M.; Goossens, A.; Leon-Reyes, A. Saponin Determination, Expression Analysis and Functional Characterization of Saponin Biosynthetic Genes in Chenopodium quinoa Leaves. Plant Sci. 2016, 250, 188–197. [Google Scholar] [CrossRef]
- Quelal, M. Análisis de la Cadena Agroproductiva de la Quinua (Chenopodium Quinoa Willd), en Las Provincias de Chimborazo e Imbabura. Bachelor’s Thesis, Ingeniera Agroindustrial, Escuela Politecnica Nacional, Quito, Ecuador, 2009. [Google Scholar]
- Villacrés, E.; Peralta, E.; Egas, L.; Mazon, N. Potencial Agroindustrial de la Quinua; Instituto Nacional Autónomo de Investigaciones Agropecuarias: Quito Ecuador, 2011; p. 32. [Google Scholar]
- Villacrés, E. La Riqueza Oculta de las Hojas de Quinua (Chenopodium quionoa Willd); INIAP, Estación Experimental Santa Catalina, Departamento de Nutrición y Calidad: Quito, Ecuador, 2016. [Google Scholar]
- Vázquez-Luna, A.; Pimentel, V.C.; Carmona, F.F.; Sobac, R.D. Quinoa Leaf as a Nutritional Alternative. Int. J. Agric. Nat. Resour. 2019, 46, 137–143. [Google Scholar] [CrossRef]
- Pichazaca, N.; Quishpilema, L.D.; Pichazaca, J.L. Innovación de la agricultura familiar kañari a través de la cadena de valor de granos para sostener la seguridad alimentaria y conservar la agro-biodiversidad local. Mem. Univ. Azuay 2018, 183–198. [Google Scholar]
- Adams, A. Lino’s Dream. Available online: https://www.esperanzaproject.com/2019/native-american-culture/linos-dream-kanari-future-traditions-ecuadorean-andes-2/ (accessed on 3 November 2020).
- Murphy, K.M.; Bazile, D.; Kellogg, J.; Rahmanian, M. Development of a Worldwide Consortium on Evolutionary Participatory Breeding in Quinoa. Front. Plant Sci. 2016, 7, 608. [Google Scholar] [CrossRef]
- Döring, T.F.; Knapp, S.; Kovacs, G.; Murphy, K.; Wolfe, M.S. Evolutionary Plant Breeding in Cereals—Into a New Era. Sustainability 2011, 3, 1944–1971. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, J.; Murphy, K. Evolutionary Participatory Quinoa Breeding for Organic Agro-Ecosystems in the US Pacific Northwest. In Farmers and Plant Breeding Current Approaches and Perspectives; Westengen, O.T., Winge, T., Eds.; Routlege Taylor & Francis Group: New York, NY, USA, 2019; p. 24. ISBN 978-1-138-58942-8. [Google Scholar]
- Murphy, K.; Lammer, D.; Lyon, S.; Carter, B.; Jones, S.S. Breeding for Organic and Low-Input Farming Systems: An Evolutionary–Participatory Breeding Method for Inbred Cereal Grains. Renew. Agric. Food Syst. 2005, 20, 48–55. [Google Scholar] [CrossRef]
- Suneson, C.A. An Evolutionary Plant Breeding Method. Agron. J. 1956, 48, 188–191. [Google Scholar] [CrossRef]
- Murphy, K.M.; Carter, A.H.; Jones, S.S. Evolutionary Breeding and Climate Change. In Genomics and Breeding for Climate-Resilient Crops: Volume 1 Concepts and Strategies; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 377–389. ISBN 978-3-642-37044-1. [Google Scholar]
- Merrick, L.F.; Lyon, S.R.; Balow, K.A.; Murphy, K.M.; Jones, S.S.; Carter, A.H. Utilization of Evolutionary Plant Breeding Increases Stability and Adaptation of Winter Wheat across Diverse Precipitation Zones. Sustainability 2020, 12, 9728. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Quinoa Line | Yield (kg ha−1) | Plant Height (cm) | Days to Harvest | Mildew Severity 1 |
|---|---|---|---|---|
| LQEP4 | 1559 | 130 | 162 | 3 |
| LQEP8 | 1708 | 125 | 160 | 3 |
| EQ26 | 1522 | 121 | 161 | 5 |
| EQ28 | 1508 | 120 | 160 | 4 |
| INIAP Tunkahuan | 1330 | 152 | 180 | 4 |
| Zone | Period | Variety | Selection Criteria | Comments |
|---|---|---|---|---|
| Alto Guanujo | 2005–2016 | “INIAP Pata de Venado” | Early, adapted to high altitude, resistant to freezing, wind, and mildew. Good flavor in soup. Used in rotation with potatoes. | Variety named “Pata de Venado” (Deer leg) (because deer are light, fast, and can resist cold, wind, and drought) |
| Middle Zone | 2005–2019 | “INIAP Tunkahuan Line ECU-6717” | Moderately early, tolerant to mildew, and resistant to lodging. The grain is white and sweet. Excellent for soups, flour, cookies. | Variety was grown in an intercrop system with corn “INIAP 111”, local corn varieties, chocho, and lentils |
| Order/Family | Functional Role |
|---|---|
| Hemiptera: Aphididae 1 | Phytophagous |
| Coleoptera: Curculionidae | Phytophagous |
| Lepidoptera: Gelechiidae | Phytophagous |
| Coleoptera: Elateridae | Phytophagous |
| Coleoptera: Latriididae | Phytophagous |
| Lepidoptera: Arctiidae | Phytophagous |
| Hymenoptera: Braconidae | Parasitoid |
| Hemiptera: Aphididae 1 | Phytophagous |
| Neuroptera: Chrysopidae | Predator |
| Lepidoptera: Noctuidae | Phytophagous |
| Order/Family | Functional Role |
|---|---|
| Coleoptera: Curculionidae 1 | Phytophagous |
| Coleoptera: Sthaphylinidae | Predator |
| Coleoptera: Tenebrionidae | Detritivore |
| Lepidoptera: Noctuidae | Phytophagous |
| Himeptera: Aphidae | Phytophagous |
| Dermaptera: Anisolabididae | Detritivore, predator |
| Coleoptera: Curculionidae 1 | Phytophagous |
| Family | Functional Role |
|---|---|
| Halictidae | Pollinator |
| Braconidae | Parasitoid |
| Chalcididae | Parasitoid |
| Ichneumunidae | Parasitoid |
| Crabronidae | Predator |
| Pteromalidae | Parasitoid |
| Bethylidae | Parasitoid, predator |
| Megaspilidae | Parasitoid |
| Diapriidae | Parasitoid |
| Figitidae | Parasitoid |
| Phase I | Phase II | ||
|---|---|---|---|
| Pedigree | Pedigree | ||
| Female | Male | Female | Male |
| Colorado 407D | 3964 | Pison | 3964 |
| Titicaca | 3964 | Japanese Strain | 3964 |
| Titicaca | Pasankalla | QQ74 | 3964 |
| Colorado 407D | Pasankalla | 23TES | 3964 |
| Titicaca | Cica | Titicaca | MisaMisa |
| Puno | Pasankalla | QQ74 | MisaMisa |
| Puno | Pasankalla | ||
| Colorado 407D | Pasankalla | ||
| Kaslaea | INIAP Tunkahuan | ||
| Titicaca | Cica | ||
| Planting | Date | Location | Elevation (m a.s.l.) | Activity |
|---|---|---|---|---|
| 2015 | Cañar Comunidad San Rafael (2°32′55.2″ S 78°57′07.0″ W) | 3073 | - Initiation of APROSANAMY and WSU collaboration | |
| Phase I | 23 May 2016 | Molobog Chico (2°36′27.252″ S 78°52′48.432″ W) | 3203 | - Kañari planting ceremony (Figure 5A) - Field preparation, plowing (Figure 5B) - Planting of barley populations with WSU students |
| La Posta (2°36′27.252″ S 78°52′48.432″ W) | 2956 | - Planting of quinoa and barley populations with WSU students (Figure 5C) | ||
| 18 March 2017 | La Posta | 2956 | - Leonardo Hinojosa and Dan Packer (WSU) visited second cycle of planting populations | |
| Phase II | 22 May 2018 | La Posta | 2956 | - Nicolas Pichazaca planted 10 F2 populations (Table 6) |
| Phase I, II and Pure Line Selection strategy | 2 February 2019 | La Posta | 2956 | - Leonardo Hinojosa and Nicolas Pichazaca increased seed from Puno/Pasankalla (Phase I) - They planted F3 population (Titicaca/CICA), (Titicaca/MisaMisa) from Phase II - They planted 60 F6 lines of (Cahuil/PI 510534) (Phase III) |
| 15 October 2019 | La Posta | 2956 | - Kevin Murphy, Nicolas Pichazaca, and Cristina Ocaña Gallegos planted 37 quinoa lines |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hinojosa, L.; Leguizamo, A.; Carpio, C.; Muñoz, D.; Mestanza, C.; Ochoa, J.; Castillo, C.; Murillo, A.; Villacréz, E.; Monar, C.; et al. Quinoa in Ecuador: Recent Advances under Global Expansion. Plants 2021, 10, 298. https://doi.org/10.3390/plants10020298
Hinojosa L, Leguizamo A, Carpio C, Muñoz D, Mestanza C, Ochoa J, Castillo C, Murillo A, Villacréz E, Monar C, et al. Quinoa in Ecuador: Recent Advances under Global Expansion. Plants. 2021; 10(2):298. https://doi.org/10.3390/plants10020298
Chicago/Turabian StyleHinojosa, Leonardo, Alex Leguizamo, Carlos Carpio, Diego Muñoz, Camilo Mestanza, José Ochoa, Carmen Castillo, Angel Murillo, Elena Villacréz, Carlos Monar, and et al. 2021. "Quinoa in Ecuador: Recent Advances under Global Expansion" Plants 10, no. 2: 298. https://doi.org/10.3390/plants10020298